Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Характеристика фагоцитирующих клетокСодержание книги
Поиск на нашем сайте
Процесс фагоцитоза начинается со стадии уз- навания и адгезии посторонней частицы (например, микробной клетки) на мембране фагоцита. При этом из фагоцита вытягиваются и сливаются друг с другом псевдоподии (ложноножки), так что микроб оказыва- ется заключенным внутри вакуоли (фагосомы), кото- рая затем сливается с лизосомой фагоцита. В результа- те микроорганизм погибает под действием биоцидных факторов фагоцита (О2, Н2О2, НСlO) и разрушается с помощью его гидролитических ферментов. Фаго- цитоз, при котором происходит гибель и разрушение поглощенных клеток, носит название завершенного. Наряду с этим при некоторых инфекциях (гонорея, туберкулез, лепра, лейшманиоз, коклюш, брюшной тиф, туляремия, бруцеллез, микозы) наблюдается не- завершенный фагоцитоз, при котором микроорганиз- мы поглощаются фагоцитами, но не погибают, а ино- гда размножаются. Это связано с тем, что некоторые микробы выделяют вещества, блокирующие процесс фагоцитоза. К ним относятся микробные токсины, ве- щества капсулы, специфические белки, лейкоцидины, токсины. Активируют фагоцитоз соли кальция, маг- ния, электролиты, комплемент, антитела. Кроме того, микроорганизмы могут избегать разрушения вслед- ствие того, что они покидают фагосому до ее слияния с лизосомой и становятся недоступными для литиче- ских ферментов. Фагоциты, кроме разрушения чужеродных клеток, еще и выполняют важную функцию антигенпредстав- ляющих клеток иммунной системы для дальнейшего развития специфического иммунного ответа. Естественные клетки-киллеры (ЕКК) — это лим- фоциты, обладающие цитотоксичностью по отноше- нию к клеткам-мишеням (опухолевым, содержащим вирусы и другие паразиты). Они не обладают фаго- цитирующей активностью, но убивают свою жертву с помощью цитотоксических веществ. Их действие неспецифично в отличие от антиген-специфических Т-киллеров (см. ниже). Среди лейкоцитов крови чело- века ЕКК составляют от 2 до 12%.
Иммунитет
Иммунитет отличается высокой специфичностью своих реакций, основанной на тонких структурно-хи- мических различиях антигенов и соответствующих им антител (иммуноглобулинов), образующихся в ответ на попадание антигена в организм [34]. По происхождению иммунитет подразделяют на врожденный (его также называют наследствен- ным или видовым) и приобретенный. Видовой им- мунитет — это невосприимчивость некоторых видов животных к болезням, поражающим другие виды. На- пример, люди не болеют кроличьим насморком, кури- ной холерой; животные не подвержены венерическим заболеваниям. Видовой иммунитет является следстви- ем длительной эволюции взаимоотношений микроба и макроорганизма и зависит от их биологических осо- бенностей, сформировавшихся в ходе естественного отбора. Степень напряженности видового иммуните- та может быть абсолютной и относительной. Напри- мер, у крыс в клетках нет рецепторов к дифтерийному токсину, поэтому они обладают абсолютной устойчи- востью к дифтерии. Для других животных восприим- чивость к некоторым инфекционным заболеваниям можно повысить, воздействуя на них какими-либо не- благоприятными факторами, например, изменяя тем- пературу тела, вводя гормоны, иммунодепрессанты, подвергая их ионизирующей радиации. В этом случае говорят об относительном врожденном иммунитете.
Приобретенный иммунитет по наследству не пе- редается. Он формируется по отношению к кон- кретному виду возбудителя и является строго спец- ифическим. Приобретенный иммунитет подразделяют на естественный и искусственный, а каждый из этих видов в свою очередь делится на активный и пассив- ный. Естественный активный иммунитет возникает после перенесенного заболевания или после инфи- цирования без клинических признаков заболевания. Естественный пассивный иммунитет — это иммуни- тет новорожденных, которые получают его от матери в период внутриутробного развития. Его продолжи- тельность невелика — около 6 месяцев. Искусствен- ный активный иммунитет возникает после активной иммунизации, т. е. введения вакцин и анатоксинов; ис- кусственный пассивный — после введения иммунных сывороток или сывороточных препаратов (иммуногло- булинов).
Антигены
Антигенами называют все вещества, которые не- сут признаки генетической чужеродности и при вве- дении в организм вызывают развитие специфических иммунологических реакций. Антигенность присуща белкам, многим полисахаридам, гликоконъюгатам (гликопротеинам, липополисахаридам), некоторым ис- кусственным высокополимерным соединениям.
Для того чтобы антиген вызвал в организме им- мунный ответ, он должен обладать достаточной моле- кулярной массой (обычно не менее 5-10 кД). Основные понятия, характеризующие антиген: антигенность (способность вызывать иммунный от- вет), иммуногенность (способность вызывать имму- нитет, т. е. невосприимчивость к инфекции) и спец- ифичность. Специфичность определяется особенностями химической структуры антигена, благодаря которым один антиген отличается от другого. Химическая груп- пировка молекулы антигена, определяющая его спец- ифичность, называется антигенной детерминантой (эпитопом). Например, антигенная специфичность белка определяется его первичной и надмолекулярны- ми структурами, а также поверхностно расположен- ными группами, которые могут служить антигенными детерминантами. В молекулах гликоконъюгатов эпи- топом часто является полисахарид. Специфичность антигена выражается в том, что он реагирует только с теми антителами и иммунными лимфоцитами, кото- рые возникли в ответ на его введение. Гаптены — это вещества, обладающие специфич- ностью, но не вызывающие иммунного ответа при введении в организм. Однако с готовыми антителами они взаимодействуют. Гаптены приобретают свойства полноценных антигенов после соединения с крупно- молекулярными веществами (белками, полисахари- дами или искусственными полиэлектролитами). Со- единение происходит за счет ковалентных связей или электростатических сил. Свойствами гаптенов могут обладать многие лекарственные препараты (амидопи- рин, хинидин, продукты распада пенициллина и др.), которые, взаимодействуя с белками организма, могут вызывать иммунный ответ — лекарственную аллер- гию. Видовая специфичность — это специфичность, благодаря которой представители одного вида отлича- ются от особей другого вида. Например, различается состав сывороточных белков у человека и животных, что может быть использовано на практике, например, в судебной медицине. Групповая специфичность обусловливает разницу среди особей одного вида. Антигены, благодаря кото- рым особи одного вида различаются между собой, на- зываются изоантигенами. К изоантигенам относятся антигены, которые содержатся в эритроцитах и опре- деляют группы крови человека. Состав молекул главного комплекса гистосовме- стимости (МНС — от англ. Major Histocompatibility Complex) уникален для каждого организма и опреде- ляет его биологическую индивидуальность, что позво- ляет отличать “свое” (гистосовместимое) от “чужого” (несовместимого). Молекулы I и II классов — это гли- копротеины, которые контролируют иммунный ответ и участвуют в реакциях цитотоксичности, осущест- вляемой Т-лимфоцитами. Молекулы I класса МНС представлены на поверхности всех ядросодержа- щих клеток, молекулы II класса — преимущественно на мембране иммунокомпетентных клеток (макрофа- гов, В-лимфоцитов, активированных Т-лимфоцитов). Гены III класса МНС кодируют отдельные компоненты системы комплемента. Типоспецифичность определяет антигенные раз- личия внутри одного вида микроорганизмов (серо- вары). Например, известно более 80 сероваров пнев- мококков, различающихся своими полисахаридными антигенами. Гетероспецифичность обусловлена наличием гетероантигенов — общих для представителей раз- ных видов антигенных детерминант. Общие антигены встречаются у весьма отдаленных видов: у человека и возбудителя чумы, вируса гриппа и других микро- организмов. Это так называемые мимикрирующие антигены. При сходстве антигенных структур микро- и макроорганизма формирование иммунного ответа нарушено, например, в миокарде человека имеются химические структуры, сходные с антигенами стреп- тококков, поэтому антитела против стрептококков спо- собны атаковать ткани сердца, что служит причиной осложнений после перенесенной скарлатины.
Антигенная структура микробной клетки пред- ставляет большой научный и практический интерес, поскольку антигены бактерий используют с целью создания вакцинных препаратов для воспроизведе- ния искусственного иммунитета и для серологической диагностики инфекционных болезней. В состав бакте- риальной клетки и вирусной частицы входят сложные комплексы веществ, обладающих антигенной актив- ностью. К ним относятся высокомолекулярные соеди- нения белковой природы, полисахариды, липополи- сахариды и т. п. Антигенными свойствами обладают органоиды клеток: жгутики, мембраны, цитоплазма, рибосомы, клеточная стенка. Токсины бактерий также являются сильными антигенами. У подвижных бактерий различают Н-антигены (жгутиковые) — термолабильные, разрушающие- ся при температуре 56-80°C, состоящие из белков, и О-антигены (соматические) — термостабильные, выдерживающие нагревание до 80-100°C, имеющие липопротеидную природу. У капсульных бактерий, например, клебсиелл, пневмококков и др. существуют полисахаридные кап- сульные антигены. Антиген вирулентности (Vi) также относится к капсульным, он описан у вирулентных видов энтеро- бактерий, например, у сальмонелл. Протективный антиген не является постоянной частью микробной клетки. Он образуется в заражен- ном организме и обладает особенно сильным иммуно- генным действием. Антигены рибосом явились основой для разработ- ки нового типа вакцины. Антигены вирусов — белки, гликоконъюгаты, ну- клеопротеиды, специфичны для вируса, или содержат компоненты клетки хозяина (липиды, углеводы). Мно- гие вирусы содержат особый антиген — гемагглюти- нин, который выявляется в реакции гемагглютинации. Эта реакция основана на способности вирусного ге- магглютинина агглютинировать эритроциты человека и животных и позволяет выявить наличие вируса в ис- следуемом субстрате. Реакция гемадсорбции основана на способности эритроцитов адсорбироваться на клетках, пораженных вирусом. Обе реакции проходят без участия антител, поэтому не являются иммунологическими. Однако ге- магглютин способен вызывать образование антител, которые могут быть обнаружены в реакции торможе- ния гемагглютинации.
Антитела
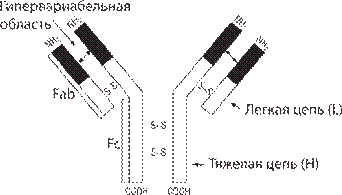
Антитела — это особые белки (гликопротеины), которые образуются в организме позвоночных жи- вотных и человека при введении антигенов и облада- ют способностью вступать с ними в специфическую связь. Антигены, связанные с антителами, обезврежи- ваются и удаляются из организма. Особое значение имеет способность антител со- единяться со специфической молекулой антигена, “уз- навать” даже самые тонкие различия в ее структуре, например, не только замену в молекуле белка одной аминокислоты на другую, но даже замену стереоизо- меров органических соединений. Специфичность ан- тител связана с различием в их химическом строении, т. е. в последовательности аминокислот полипептид- ных цепей, составляющих их структуру. Наряду с тон- кими различиями в структуре антител, определяющи- ми их иммунологическую специфичность, антитела имеют общие характерные особенности строения. Антитела имеют глобулярную структуру и назы- ваются иммуноглобулинами. Основу структуры любо- го антитела (рис. 79) составляет комплекс из четырех полипептидных цепей — двух одинаковых тяжелых и двух одинаковых легких. Тяжелые цепи обознача- ют буквой Н (heavy — тяжелый), а легкие — буквой L (light — легкий). Тяжелые и легкие цепи удержива- ются вместе дисульфидными мостиками. Они уложе- ны таким образом, что на поверхности образующейся структуры возникают два одинаковых участка, ко- торые обозначаются Fab. Эти участки содержат цен- тры связывания антигена. Третий участок Fc содер- жит структуры, обеспечивающие связывание антител с определенными клетками, несущими на своей по- верхности рецепторы Fc-фрагмента, например, лей- коцитами, тучными клетками. Центры связывания антигена (активные центры) соответствуют эпитопам антигена (подходят друг другу как замок и ключ). Взаимодействие антигена с антителом осуществляет- ся за счет электростатических, гидрофобных взаимо- действий и сил Ван-дер-Ваальса. Молекулы полных антител имеют как минимум два центра связывания с антигеном. Антитела, имеющие только один центр связывания, называют неполными.
Рис. 79. Структура мономера молекулы иммуноглобу- лина [35, 36].
Как тяжелые, так и легкие цепи в своей структуре имеют две категории областей: вариабельные (V) и по- стоянные (С) по содержанию составляющих их ами- нокислот. Вариабельная область связана со специфич- ностью антител. Она находится в участке Fab, тогда как постоянная область — в участке Fc. Вторичная структура иммуноглобулинов — а-спираль, переме- жающаяся сложными β-структурами — “клубками”, возникающими при сшивании аминокислотных остат- ков каждой цепи. Эти клубки называют доменами, они находятся на тяжелых и легких цепях вариабельных и константных участков. Активные центры антител формируются доменами вариабельных участков, ко- торые располагаются в гипервариабельных областях тяжелых и легких цепей.
Основные свойства иммуноглобулинов
Иммуноглобулины представляют собой гетеро- генную группу белков, их гетерогенность связана с су- ществованием разных типов тяжелых и легких цепей. У человека имеется два типа легких цепей: κ и λ, (кап- па и лямбда) и 5 классов тяжелых цепей α, γ, μ, δ, ε. В соответствии с классами тяжелых цепей иммуногло- булины подразделяют на 5 классов (IgG, IgM, IgA, IgD, IgE), различающихся по своим физико-химическим свойствам и биологической активности (табл. 40). IgM — пентамер из 5 субъединиц, имеет 10 цен- тров связывания с антигеном. Филогенетически наибо- лее древний, наиболее ранний, обнаруживаемый при первичном попадании антигена в организм, основной класс, синтезируемый у новорожденных и младенцев. IgG составляют до 75% всех иммуноглобулинов, это основной класс, защищающий организм от бак- терий, вирусов, токсинов и обеспечивающий форми- рование пассивного иммунитета у плода, поскольку только эти иммуноглобулины способны преодолевать плацентарный барьер. IgA существует в двух видах — сывороточном и секреторном. Большая часть IgA — это секреторные иммуноглобулины (slgA), у которых два или три моно- мера соединены секреторным фрагментом, защища- ющим иммуноглобулин от разрушения ферментами. slgA присутствуют в слюне, слезах, молоке, секрети- руются на поверхности эпителия и слизистых оболо- чек, усиливая их защитные функции. IgE специфически взаимодействуют с тучными клетками и базофильными лейкоцитами и принимают участие в развитии аллергических реакций. Их защитные функции направлены в основном против гельминтов. IgD функционируют как мембранные рецепторы антигена на В-лимфоцитах. Кроме различных классов (изотипов) иммуногло- булинов, между ними существуют аллотипические и идиотипические различия. Аллотипы (маркеры константной области) генетически детерминированы и наследуются. Идиотины определяют индивидуаль- ную характеристику каждого антитела. Идиотипиче- ские маркеры (антигенные детерминанты) локализу- ются в гипервариабельных областях и соответствуют антигенсвязывающим участкам антител. Все молеку- лы иммуноглобулинов, продуцируемые одним клоном лимфоцитов, несут один и тот же идиотип и называ- ются моноклональными антителами. Источником разнообразия идиотипов служат мутации ДНК, коди- рующей вариабельные участки, нематричный синтез ДНК между рекомбинирующими сегментами и сдвиг точки соединения сегментов на несколько нуклеоти- дов. Иммунитет, опосредованный антителами, находя- щимися в плазме крови и других жидкостях организма, называется гуморальным. Его механизм связан со спо- собностью антител нейтрализовать возбудитель и его токсины, посредством опсонизации, антитоксического действия, активации комплемента и других воздей- ствий на инфекционный агент. Опсонизация проис- ходит путем связывания антител с поверхностью ми- кробной клетки. Такой комплекс активно поглощается фагоцитом при взаимодействии Fc-фрагмента антите- ла с соответствующим Fc-рецептором фагоцита. Анти- тела способны взаимодействовать с рецепторами кле- ток, связывающими бактерии или вирусы, препятствуя их адгезии и проникновению в клетки организма-хо- зяина. Кроме того, антитела обладают каталитической активностью, действуя как гидролазы и оксидоредук- тазы, принимают участие в нейтрализации инфекци- онного агента.
Иммунная система
Иммунная система представляет собою совокуп- ность лимфоидных органов и лимфоидных клеток, которые распространяются по всему телу организ- ма и связаны системой кровообращения, лимфотока и единой системой иммунорегуляции. Иммунная си- стема обладает уникальной способностью выраба- тывать молекулы антител, специфичные для каждого антигена. Органы иммунной системы подразделяются на первичные — центральные (костный мозг и тимус) и вторичные — периферические (селезенка, лимфати- ческие узлы, скопления лимфоидной ткани). Первичные — центральные органы иммунной системы обеспечивают ее самообновление. В них происходят процессы пролиферации клеток — пред- шественников, их дифференцировка и созревание, в результате чего они превращаются в иммунокомпе- тентные клетки, выходят в циркуляцию и заселяют пе- риферические органы иммунной системы. Костный мозг — место образования полипотент- ной стволовой клетки, которая дает начало разным росткам кроветворения, в том числе миеломоноцитар- ному и лимфоцитарному. В костном мозге образуются цитокины; у млекопитающих это возможное место со- зревания В-лимфоцитов (см. ниже). Тимус (вилочковая железа) — место созревания и дифференциация Т-лимфоцитов. Тимус активно вырабатывает лимфоциты в период эмбриогенеза, достигает максимального размера у большинства по- звоночных вскоре после рождения, затем постепенно инволюционирует. У человека самая высокая продук- ция Т-лимфоцитов сохраняется до двух лет жизни, за- тем быстро падает. Однако количество Т-лимфоцитов сохраняется на достаточном уровне, т. к. это долгожи- вущие клетки, кроме того, они способны пролимфери- ровать в ответ на встречу со специфичным антигеном. В процессе созревания и дифференциации предше- ственники Т-лимфоцитов из костного мозга поступа- ют в корковый слой тимуса, постепенно мигрируют внутрь и приобретают свои маркеры, контактируя с клетками тимуса и продуцируемыми ими медиатора- ми (специфическими пептидами) и цитокинами. В тимусе происходит элиминация потенциаль- но аутореактивных клеток. Основная функция зре- лых Т-лимфоцитов — распознавание чужеродных антигенов, но не антигенов собственного организ- ма (аутоантигенов). Элиминация состоит в том, что Т-лимфоциты, имеющие рецепторы для аутоантиге- нов (их в популяции 95-98%), получают сигнал для апоптоза, а Т-клетки, обладающие рецепторами для чужеродных антигенов — сигнал для пролиферации. Апоптоз (программированная гибель) — это процесс распада клетки на отдельные фрагменты, которые мо- гут быть использованы для построения других клеток. Вторичные — периферические органы иммунной системы — это место встречи иммунокомпетентных клеток с антигеном, его распознавания и развития специфического иммунного ответа (взаимодействия иммунокомпетентных клеток и синтеза иммуноглобу- линов). Лимфатические узлы распространены по всему телу организма. Один лимфоузел имеет массу около 1 г и содержит приблизительно 2´109 лимфоцитов. Каждый час из него выходит в лимфу количество лим- фоцитов, равное его утроенному весу. Он отфильтро- вывает микробные клетки и другие частицы, в нем раз- вивается иммунный ответ на антигены, попадающие в лимфу. В селезенке развивается иммунный ответ на анти- гены, попадающие в кровь. Она удаляет из крови чуже- родные частицы, а также состарившиеся и поврежден- ные эритроциты. При удалении селезенки ее функцию берут на себя лимфоидные органы, у таких пациентов иммунитет ослаблен. Лимфоидная ткань ассоциирована со слизи- стой оболочкой кишечника, глотки, дыхательных пу- тей, мочеполового тракта. Ее функция — активация В-лимфоцитов, продуцирующих иммуноглобулины классов А и Е.
Иммунокомпетентные клетки
Иммунокомпетентные клетки — это клетки, при- нимающие участие в иммунном ответе. Они постоян- но циркулируют между кровью, лимфой и лимфоид- ными органами, чтобы обеспечить встречу со “своим” антигеном, т. к. каждый антиген распознается лишь небольшой частью популяции лимфоцитов. Лимфоци- ты — единственные клетки, способные распознавать антиген и отвечать на контакт с ним. Индивидуальные лимфоциты специализированы: они способны (ком- митированы) отвечать лишь на определенную группу структурно сходных антигенов. Эта коммитирован- ность существует еще до первого контакта с антигеном и выражается в наличии у лимфоцита мембранных ре- цепторов, специфичных для детерминант определен- ного антигена. Лимфоциты являются исключительно неоднородной популяцией клеток: считают, что число рецепторов лимфоцитов с различными антигенсвязы- вающими центрами составляет не менее 106. Кроме того, лимфоциты различаются по их функциям в про- цессе иммунного ответа и другим физиологическим особенностям. В-лимфоциты получили свое название потому, что исследования, проведенные на птицах, показа- ли, что местом их дифференциации является особый лимфоидный орган — бурса (сумка) Фабрициуса. У млекопитающих это лимфоидный орган, о лока- лизации которого пока нет единого мнения (костный мозг, лимфоидная ткань слизистой оболочки кишеч- ника). В-лимфоциты в ходе иммунного ответа диффе- ренцируются в клетки, продуцирующие иммуногло- булины. Они распознают антигенные детерминанты с помощью рецепторов, представляющих иммуногло- булины классов D или М. Кроме того, они обладают другими рецепторами, для распознавания сигналов в процессе иммунного ответа. Они способны распоз- навать антиген в растворе и связывать белковые, по- лисахаридные и липопротеиновые антигены. Срок жизни большей части популяции В-лимфоцитов (около 85% ) составляет не более 10 дней, около 14% живут 4-6 недель, около 1% жи- вут десятилетиями, это клетки иммунологической памяти. Т-лимфоциты — основные клетки иммунологи- ческой памяти, они различаются по своим функци- ям, дифференцируются по поверхностным маркерам, определяемым серологическими методами. Т-эффекторы СД8 + продуцируют многочис- ленные цитокины, которые способствуют их диф- ференциации и пролиферации, а также активируют В-лимфоциты и макрофаги. Цитотоксические лимфоциты (CTL, Т-киллеры) имеют маркер СД8+, они появляются в результате акти- вации и пролиферации Т-эффекторов под воздействи- ем антигена. При размножении паразита (микобакте- рии туберкулеза, грибы, простейшие, вирусы) внутри клетки хозяина его антигены образуют комплекс с ан- тигенами МНС I класса, который распознается рецеп- торами CTL. В ответ CTL активируются, т. е. приоб- ретают способность убивать инфицированную клетку с помощью пресинтезированных цитотоксинов: фраг- ментинов и перфоринов. Первые индуцируют апоптоз клетки-мишени, вторые представляют собою белки, образующие поры, нарушающие проницаемость мем- браны клетки-мишени и прокладывающие путь фраг- ментинам. Контакт CTL и клетки-мишени непродол- жителен, после чего CTL движется к новой жертве. Клетки с маркером СД8+ обладают и супрессорной активностью, т. е. регулируют интенсивность иммун- ного ответа и предотвращают развитие аутоиммунных реакций. Регуляторные СД4+ лимфоциты или Т-хелперы (ТН) подразделяют на две группы: ТН-1 (воспали- тельные) активируют макрофаги и принимают уча- стие в реакциях гиперчувствительности замедленного типа; ТН-2 активируют В-лимфоциты в процессе им- мунного ответа. Макрофаги обеспечивают неспецифическую за- щиту за счет фагоцитоза и играют важную роль в раз- витии иммунного ответа как антигенпроцессирующие и антигенпредставляющие клетки. Они происходят из стволовой клетки костного мозга, проходят стадии циркулирующих в крови промоноцита и моноцита и мигрируют в ткани (стадия гистиоцита или ткане- вого макрофага). Они несут на своей поверхности рецепторы (маннозы, липополисахаридов), необхо- димые для захвата микробных клеток, а также рецеп- торы Fc-области иммуноглобулинов и комплемента, необходимые для осуществления реакций с участием антител. Макрофаги и цитотоксические лимфоциты осу- ществляют клеточный иммунный ответ (реакции гиперчувствительности замедленного типа, уничто- жение собственных инфицированных и опухолевых клеток). Дендритные клетки происходят из костного мозга и заселяют нелимфоидные ткани. Дендритные клетки эпидермиса кожи и в слизистых оболочек дыхательных путей называются клетками Лангерганса. Участвуют в иммунном ответе как антигенпредставляющие клет- ки. Сходны с макрофагами, но фагоцитирующей ак- тивностью обладают только их незрелые формы.
Цитокины
Цитокины — это полипептиды, продуцируемые иммунокомпетентными клетками и в то же время яв- ляющиеся эффекторами этих клеток, действующими на их метаболизм, пролиферацию и секрецию. Их ос- новные функции: контроль пролиферации клеток, кон- троль воспалительных реакций, регуляция клеточного и гуморального иммунного ответа, противовирусная активность и цитотоксичность. Многие из них полифункциональны (табл. 41). Нарушение активности или синтеза цитокинов приво- дит к заболеваниям [34]. Некоторые цитокины полу- чают методами генетической инженерии и используют как лечебные препараты.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-05; просмотров: 113; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.231.199 (0.013 с.) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||