Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Проксимальный извитой каналецСодержание книги
Поиск на нашем сайте
В проксимальных извитых канальцах обратному всасыванию подвергается 65-75 % воды и натрия, которые содержатся в протекающем по ним фильтрате. Отметим, что реабсорбируется изотоническая жидкость, т. е. содержащая пропорциональное
Рис. 31-1. Схема строения нефрона. (Из: Ganong W. F. Review of Medical Physiology, 14th ed. Appleton & Lange, 1989; с разрешения.) количество воды и натрия (рис. 31-2). В процессе реабсорбции большинство веществ проникают вначале через люминальную (т. е. обращенную в просвет канальца) поверхность клеточной мембраны, затем через базолатеральную поверхность перемещаются в почечный интерстиций и уже оттуда поступают в околоканальцевый капилляр. Главная функция проксималъного канальца состоит в реабсорбции натрия. При участии мембрано-связанной НаУЮ-зависимой АТФ-азы натрий активно перемещается из клеток проксимального канальца в почечный интерстиций к околоканаль- цевым капиллярам (рис. 31-3). Внутриклеточная концентрация натрия снижается, и он начинает пассивно поступать по градиенту концентрации из канальцевой жидкости в эпителиальные клетки. В наиболее проксимальном отделе извитого канальца реабсорбция натрия усиливается под воздействием ангиотензина II и норадреналина. До-фамин, напротив, уменьшает реабсорбцию натрия в проксимальных отделах. Реабсорбция Na+ сопряжена с реабсорбцией других растворенных веществ и секрецией H+ (рис. 31-3). Специфический белок-переносчик ТАБЛИЦА 31 -1. Функциональное подразделение нефрона^
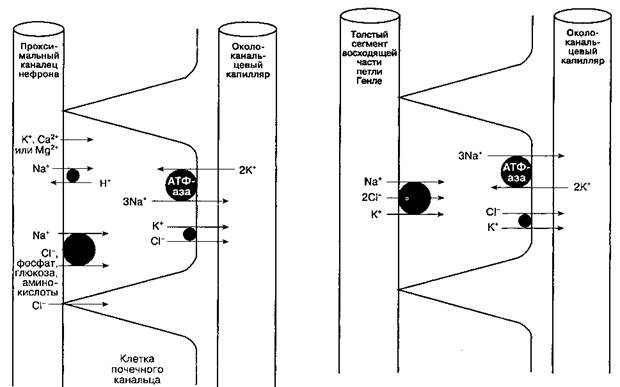
1 С изменениями. Из: Rose В. D. Clinical Physiology of Acide Base and Electrolite Disordsers, 3rd ed. McGraw-Hill,1989. 2 Частично увеличивается под действием ангиотензина II. 3 Угнетается паратиреоидным гормоном. 4 Частично опосредовано альдостероном. 5 Усиливается паратиреоидным гормоном. 6 Опосредовано антидиуретическим гормоном. 7 Угнетается предсердным натрийуретическим пептидом. при посредстве низкой внутриклеточной концентрации Na^ осуществляет транспорт фосфатов, глюкозы и аминокислот. В результате активности №+/К+-зависимой АТФ-азы (обменивающей 3 Na^ на-2 K+) уменьшается внутриклеточный положительный заряд, что облегчает реабсорбцию других катионов (K+, Ca2+, Mg2+). Именно так Ка+/К+-за-висимая АТФ-аза, расположенная на базолате-ральной поверхности эпителиальной клетки канальца, играет роль источника энергии для реабсорбции большинства растворенных в фильтрате веществ. Реабсорбция Na+ на люминальной поверхности клеточной мембраны сопряжена с секрецией H+. За счет работы этого механизма реабсорбируется до 90 % фильтруемых в клубочках ионов бикарбоната (рис. 30-2). В отличие от других растворенных веществ, хлориды способны проходить через узкие щели между эпителиальными клетками канальцев. Поэтому их реабсорб-ция осуществляется пассивно по градиенту концентрации. Кроме того, за счет функционирования КуСГ-транспортера, который перемещает оба иона на капиллярную поверхность клеточной мембраны (рис. 31-3), происходит и активный механизм реабсорбции хлоридов. В проксимальных канальцах секретируются органические катионы и анионы. Ряд органических веществ-катионов (креатинин, циметидин и хини-дин), конкурируя за один и тот же насосный механизм переноса, способны препятствовать экскреции друг друга. Доказано, что общие механизмы секреции имеют такие органические анионы, как ураты, кетоновые кислоты, пенициллины, цефа-лоспорины, диуретики, салицилаты и большая часть рентгеноконтрастных препаратов. Оба насоса, вероятно, играют основную роль в элиминации различных циркулирующих токсинов. Фильтруемые низкомолекулярные белки в норме реабсорби-руются клетками проксимальных канальцев и подвергаются в них метаболической деградации. Петля Генле Петля Генле состоит из нисходящей и восходящей частей. Тонкий сегмент нисходящей части является продолжением проксимального канальца и опускается из коркового вещества почки в мозговое. В мозговом веществе тонкий сегмент загибается, делая U-образный разворот, и поднимается в корковое вещество уже как восходящая часть петли Генле. В восходящей части выделяют различающиеся в функциональном отношении тонкий сегмент, толстый медуллярный сегмент и толстый кортикальный сегмент (рис. 31-1). У нефронов, клубочки которых расположенные вблизи мозго-
Рис. 31-2. Реабсорбция натрия в нефроне. Указанное количество реабсорбированного натрия в каждом отделе нефро-на соответствует проценту от профильтровавшейся натриевой нагрузки. (Из: Cogan M. G. Fluid and Electrolytes: Physiology and Pathophysiology, 14th ed. Appleton & Lange, 1991; с разрешения.) вого вещества (юкстамедуллярные нефроны), петля Генле более длинная, чем у нефронов, клубочки которых лежат ближе к наружной поверхности почки (корковые нефроны). У корковых нефронов с короткой петлей отсутствует тонкий сегмент восходящей части. Корковых нефронов в 7 раз больше, чем юкстамедуллярных. Петля Генле поддерживает гипертоничность интерстициальной жидкости мозгового вещества, а также опосредованно обеспечивает процесс концентрирования мочи в собирательных трубочках. В норме до 25-35 % фильтрата, образующегося в капсуле Боумена, достигает петли Генле. В ней реабсорбируется 15-20% фильтруемого натрия. За исключением толстого сегмента восходящей части, реабсорбция растворенных веществ и воды в петле Генле происходит пассивно — по градиенту концентрации и осмотическому градиенту соответственно. В толстом сегменте восходящей части Na" и СГ реабсорбируются в большей степени, чем вода; более того, в этой части нефрона реабсорбция Na" непосредственно сопряжена с реабсорбцией K+ и СГ (рис. 31-4), и концентрация СГ в канальцевой жидкости является фактором, ограничивающим скорость реабсорбции. Активная реабсорбция Na+ осуществляется Ка"УК+-зависимой АТФ-азой капиллярной поверхности эпителиальных клеток. В отличие от нисходящей части и тонкого сегмента восходящей части петли Генле, толстый сегмент восходящей части непроницаем для воды. Поэтому оттекающая из петли Генле канальцевая жидкость гипотонична (100-200 мОсм/л), а окружающая петлю Генле интерстициальная жидкость гипертонична. Механизм противоточного умножения работает таким образом, что гипертонич-ностъ каналъцевой жидкости и окружающего uh-терстиция значительно нарастает по мере углубления в мозговое вещество почки (рис. 31-5). Концентрация мочевины в мозговом веществе становится высокой, что существенно влияет на его гипертоничность. Структуры механизма противоточного умножения включают петлю Генле, кортикальные и медуллярные собирательные трубочки и сопровождающие их капилляры (vasa recta), Толстый сегмент восходящей части петли Генле играет важную роль в реабсорбции Ca2+ и Mg2+. В этом участке нефрона паратиреоидный гормон может увеличивать реабсорбцию кальция. Дистальный извитой каналец В дистальный извитой каналец поступает гипото-ничная жидкость из петли Генле. Как правило, состав и объем канальцевой жидкости в этом отделе нефрона изменяется значительно. В отличие от более проксимальных отделов нефрона, клетки эпителия дистальных канальцев плотно прилежат друг к другу, что делает их относительно непроницаемыми для воды и натрия, за счет чего поддерживаются градиенты концентраций, сформированные петлей Генле. В дистальных канальцах реабсорбируется 5 % фильтруемого в клубочках натрия. Реабсорбция натрия обеспечивается за счет функционирования КауК+-зависимой АТФ-азы на обращенной к капилляру поверхности клеток, а на люминальной мембране осуществляется Na^/СГ-переносчиком. Реабсорбция натрия в дистальном канальце прямо пропорциональна его поступлению. Дистальные канальцы — основное место реабсорбции кальция; этот процесс регулируется паратиреоидным гормоном и витамином D. Наиболее дистальный отдел канальца называют соединяющим сегментом. В этом участке, как и в более проксимальных, происходит опосредованная гормонами реабсорбция кальция, но в отличие от них здесь имеет место еще и регулируемая альдостероном реабсорбция натрия. Собирательная трубочка В собирательной трубочке реабсорбируется 5-7 % фильтруемого в клубочках натрия, в ней выделяют кортикальную и медуллярную части. А. Кортикальная часть собирательной трубочки. Этот сегмент нефрона состоит из двух типов клеток: 1) главных клеток, секретирующих калий и участвующих в опосредованной альдостероном реабсорбции натрия; 2) вставочных клеток, ответственных за регуляцию кислотно-основного равновесия. Главные клетки реабсорбируют натрий с помощью электрогенной помпы, поэтому для поддержания электронейтральности должна происходить либо реабсорбция СГ, либо секреция K+.
Рис. 31-3. Реабсорбция фосфатов, глюкозы, аминокислот и электролитов в проксимальном извитом канальце. Отметим, что №+/Кт-зависимая АТФ-аза, поддерживая низкую концентрацию натрия внутри клеток канальце-вого эпителия, обеспечивает энергетические потребности для реабсорбции катионов Рис. 31-4. Реабсорбция натрия и хлора в толстом сегменте восходящей части петли Генле. Белок-переносчик (расположенный на люминальной поверхности клетки петли Генле) функционирует, только если заняты все четыре места связывания. Концентрация хлора в канальцевой жидкости является фактором, ограничивающем скорость реабсорбции
Рис. 31-5. Механизм противоточного умножения, обу слов лен ный различиями в проницаемости и трансмембранном транспорте, которые существуют между нисходящей и восходящей частями петли Генле. Нисходящая часть и тонкий сегмент восходящей части петли Генле проницаемы для воды, Na+, Cl и мочевины. Толстый сегмент восходящей части петли Генле непроницаем для воды и мочевины, в нем активно реабсорбируется Na+ и Cl. Эти факторы приводят к возникновению осмотического градиента. На рисунке представлен осмотический градиент между нисходящей и восходящей частями петли Генле, постепенно увеличивающийся от О до 200 мОсм/кг. Заметим, что в петле Генле по ходу тока канальцевой жидкости градиент не изменяется, в то время как осмоляльность увеличивается по мере приближения к точке перегиба. (По: Pitts R. F. Physiology of the Kidney and Body Fluids, 3rd ed. Year Book, 1974.) Увеличение концентрации внутриклеточного калия способствует его секреции. В данном отделе нефрона альдостерон повышает активность NaV К+-зависимой АТФ-азы, увеличивая количество открытых K+- и Na^-каналов в люминальной мембране. Альдостерон также повышает активность ГГ-секретирующей АТФ-азы на люминальной поверхности вставочных клеток (рис. 31-6). Кроме того, во вставочных клетках люминальная Ю/Н+-зависимая АТФ-аза реабсорбирует K+ и секрети-рует ГГ. Некоторые вставочные клетки способны секретировать бикарбонат при большой щелочной нагрузке. Б. Медуллярная часть собирательной трубочки — основная мишень антидиуретического гормона (АДГ, синоним — аргинин-вазопрессин). АД Г активирует аденилатциклазу через У2-рецеп-торы (активация V1-рецепторов повышает сосудистое сопротивление за счет усиления метаболизма фосфатидилинозитола). Проницаемость люминальной мембраны для воды полностью зависит от присутствия АДГ (гл. 28). Дегидратация приводит к повышению секреции АДГ, в результате действия которого люминальная мембрана становится проницаемой для воды. По осмотическому градиенту вода удаляется из канальцевой жидкости, и образуется концентрированная моча (до 1400 мОсм/л). Напротив, адекватная гидратация подавляет секрецию АДГ; в этом случае осмоляльность канальцевой жидкости в медуллярной части собирательных трубочек не изменяется, и она остается гипотоничной (100-200 мОсм/л). В стенке медуллярной части собирательных трубочек содержатся основные и вставочные клетки, причем последние преобладают. Кроме того, эта часть нефрона отвечает за подкисление мочи; образующиеся в этих клетках ионы водорода выделяются в виде титруемых кислот (фосфаты) и ионов аммония (гл. 30). Предсердный натрийуретический пептид способен снижать реабсорбцию Na" в собирательных трубочках. В. Роль собирательных трубочек в поддержании гипертоничности мозгового вещества. Кортикальная часть собирательных трубочек полностью проницаема для мочевины, тогда как медуллярная часть в норме для нее непроницаема. Этим разли-
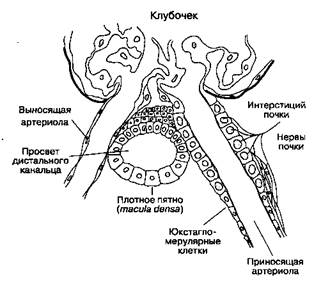
Рис. 31-6. Кортикальная часть собирательной трубочки: секреция ионов водорода, реабсорбция бикарбоната и калия чием в проницаемости практически наполовину обусловлена гипертоничность интерстициальной жидкости мозгового вещества. В присутствии АДГ значительно увеличивается проницаемость для мочевины наиболее дистальных отделов медуллярной части собирательных трубочек. Таким образом, при секреции АДГ вода покидает собирательные трубочки, и моча становится более концентрированной. Затем мочевина диффундирует в интерстициальную жидкость мозгового вещества, увеличивая ее осмоляльность. Юкстагломерулярный аппарат Этот небольшой орган, имеющийся в каждом неф-роне, состоит из специализированного сегмента приносящей артериолы, в стенке которой содержатся юкстагломерулярные клетки, и конечной части толстого сегмента восходящей части петли Генле — плотного пятна (macula densa\ рис. 31-7). Юкстагломерулярные клетки содержат фермент ренин и иннервируются симпатическими нервными волокнами. Факторы, влияющие на высвобождение ренина: 1) стимуляция fii-адренорецепто-ров; 2) давление в приносящей артериоле (гл. 28); и 3) концентрация ионов хлора в жидкости,
Рис. 31-7. Юкстагломерулярный аппарат. (С изменениями. Из: Ganong W. F. Review of Medical Physiology, 14th ed. Appleton & Lange, 1989.) протекающей через плотное пятно. Высвобождаемый в кровоток ренин воздействует на продуцируемый печенью белок — ангиотензиноген, в результате чего образуется ангиотензин I. Под воздействием ангиотензинпревращающего фермента (АПФ) физиологически инертный декапептид ангиотензин I быстро превращается в октапептид ангиотензин II. Процесс идет главным образом в сосудах легких. Ангиотензин II играет важную роль в регуляции АД (гл. 19) и секреции альдостерона (гл. 28). В клетках прокси-мальных канальцев содержатся АПФ, а также рецепторы ангиотензина IL Кроме того, внутрипо-чечное образование ангиотензина II увеличивает реабсорбцию натрия в проксимальных канальцах. Вне почек ренин и ангиотензин II вырабатываются в эндотелии сосудов, в надпочечниках и в ткани головного мозга. Почечное кровообращение Функция почек находится в тесной зависимости от кровотока в них. Почки — это единственный орган, в котором потребление кислорода зависит от кровотока; в других органах такой зависимости нет. На почечный кровоток (в обеих почках) приходится 20-25 % сердечного выброса. Кровь в почку поступает по почечной артерии, отходящей от аорты. В воротах почки почечная артерия разветвляется на междолевые артерии, от которых на границе коркового и мозгового вещества отходят дуговые
Рис. 31-8. Артерии почки. (С разрешения. Из: Leaf A., Cotran R. S. RenalPatophysiology. Oxford Univ. Press, 1976.) артерии (рис. 31-8). Дуговые артерии делятся на междольковые артерии, которые отдают к каждому клубочку по одной приносящей артериоле. От каждого клубочка отходит единственная выносящая артериола, по которой осуществляется отток крови. В свою очередь выносящие артериолы распадаются на капилляры, оплетающие почечные канальцы. В отличие от капилляров клубочков, где происходит фильтрация, в околоканальцевых капиллярах осуществляется реабсорбция. Венулы, собирающие кровь из околоканальцевого капиллярного сплетения, через почечную вену возвращают кровь в нижнюю полую вену.
|
|||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-20; просмотров: 794; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.43.200 (0.009 с.) |