
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Организация клеточного цикла дрожжейСодержание книги
Поиск на нашем сайте
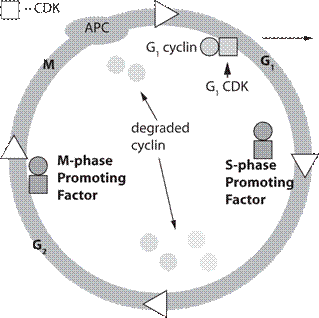
Наиболее исследован клеточный цикл у дрожжей Saccharomyces cerevisiae., хотя многие положения яв- ляются общими для всех эукариотических клеток (рис. 21). Клеточный цикл эукариотов разделен на четыре фазы: G1, S, G2, М. Синтез ядерной ДНК занимает лишь некоторую часть интерфазы — период, называемый S-фазой клеточного цикла. Между концом М-фазы и на- чалом синтеза ДНК имеется интервал, известный как фаза G 1 (от англ, gap — промежуток). Другой интервал, называемый фазой G 2, отделяет конец синтеза ДНК от начала М-фазы. Таким образом, интерфаза состоит из последовательности фаз: G1, S, G2 и обычно занимает не меньше 90% всего времени клеточного цикла. В кле- точном цикле выделяют три особых критических точки: в фазе G1, в фазе S и в фазах G2+M (G2/M -точка), ко- торые контролируются внутриклеточными процессами. Продолжительность фаз клеточного цикла неодинакова; в среднем фаза G1 составляет 30-40% от всей продол- жительности клеточного цикла, фаза S — 30-50%, фаза G2 — 10-20% и фаза М 5-10%. Эти параметры могут достаточно сильно варьироваться в зависимости от био- логического вида, но достаточно неизменны для кон- кретного типа клеток. Существует особая критическая точка в фазе G1 названная точкой старта (Start). При прохождении этой точки (для высших эукариотов — точка рестрикции) в клетке происходят изменения, по- сле которых она должна уже пройти все последующие этапы клеточного цикла. Такое детерминирование кле- точного цикла предполагает внутриклеточный контроль завершения каждой из фаз клеточного цикла (Hartwell L.H., 1974). Иными словами, после точки старта клетка детерминирована в завершении цикла и выходы из него невозможны (но не остановки) в любой другой фазе (S, G2, М). Такое детерминирование клеточного цикла предполагает внутриклеточный контроль завершения каждой из фаз клеточного цикла. События, происходя- щие, например, в фазе G2, не могут разворачиваться при прохождении фазы S, и наоборот. Если после окончания клеточного цикла не имеет- ся условий для вхождения в новый цикл, клетки пере- ходят в состояние GO. По своему определению это со- стояние просто характеризует клетки, не включенные в клеточный цикл. В этом состоянии могут находить- ся клетки, которые подготавливаются к входу в кле- точный цикл (например, увеличивают свой размер до критического, необходимого для прохождения точ- ки Старта), в этом случае тестируется (как бы) времен- ное увеличение фазы G1. Иногда, такими добавочными условиями для вхождения в клеточный цикл, является синтез не конституционных ферментных систем, не- обходимых для специфического субстрата (например, для роста дрожжей на н-алканах необходима индукция системы Р-450 (Галынкин В. А. и др., 1990)). С другой стороны, в состоянии GO находятся и клетки, осущест- вляющие синтез целевого продукта (например, био- синтез антибиотиков), в этом случае мы имеем дело с физиологической дифференцировкой. В состоянии GO находятся клетки, осуществляющие морфологиче- скую дифференцировку (например, образование спор). В некоторых случаях морфологическая и физиологи- ческая дифференцировка совпадают (синтез некото- рых антибиотиков и переход к споруляции совпадает по времени). Наконец, морфологически дифференци- рованые клетки могут войти в клеточный цикл в этом статусе. В этом случае наблюдается рост не отдельных клеток, а в виде мицелия, что также приводит к средне- му увеличению времени деления клеток. По существу, из GO состояния клетки либо возвращаются в клеточ- ный цикл (иногда в новом морфологическом статусе), либо переходят к программированной клеточной гибе- ли (автолизная программа). Для дрожжей Saccharomyces cerevisiae (рис. 21) один из генов, контролирующих прохождение точ- ки старта, был выявлен и назван cdc28 (аналог cdc2). Белок CDC28, кодируемый этим геном, представляет собой протеинкиназу, но для функционирования это- го белка требуется наличие другого белка, названного циклином. Именно циклин и является тем белком, ко- торый периодически синтезируется в ходе клеточного цикла и исчезает к его окончанию (Э. У. Мюрей, 1991). Дальнейшие исследования показали, что уместно говорить не об одном циклине, а о целом семействе циклинов. Были найдены три группы циклинов: ци- клины фазы Gi (GI cyclins), контролирующие вхож- дение в цикл (точка старта); циклины фазы S (S-phase cyclins), контролирующие прохождение фазы S; ци- клины фазы М (M-phase cyclins), контролирующие завершение цикла. Причем циклины, ответственые за каждую из фаз цикла, синтезируются в своей фазе, а затем ферментативно деградируют (распадаются) (Dirick L. 1995, Nasmyth К., 1996). Было показано, что фазы цикла инициируются растворимыми цитоплазма- тическими факторами. Активатор М-фазы был назван М-стимулирующим фактором (М-phase- promoting factor, MPF), а активатор S-фазы назван S-стимулирующим фактором (S-phase-promoting factor). MPF имеет универсальное значение для эукари- отических клеток и в эволюции высококонсервативен. Механизм регуляции цикла построенный на ак- тивации циклинзависимых протеинкиназ (cyclin- dependent kinases — CDK), выполняется для всех эукариотов, но его детали сильно различаются для конкретного типа эукариотической клетки. На рисунке 22 показано бесполое размножение дрожжей Saccharomyces cerevisiae с одновременным прохождением биохимических процессов и изменени- ем генетического статуса клетки. Половое размножение у грибов, связанный с ним процесс смены ядерных фаз, строение половых орга- нов — всё это отличается у разных групп грибов. Су- щественнейшие моменты полового процесса у грибов: плазмогамия, кариогамия и мейоз. Соответственно, гриб может находиться в гаплоидной или диплоидной стадиях. Плазмогамия и кариогамия у сумчатых и ба- зидиальных грибов не совпадают во времени, в резуль- тате чего после плазмогамии у них возникает особая стадия дикариотичного мицелия, когда гаплоидные ядра попарно ассоциированы, сближены, но не сли- лись и образуют дикарион. Ядра дикариона обычно синхронно делятся с параллельным расположением осей веретен деления. В определенный момент цикла развития они сливаются, образуя диплоидное ядро, ко- торое затем делится редукционно. Смена ядерных фаз будет подробно рассмотрена при описании отделов и классов грибов.
Рис. 21. Показаны четыре фазы: G, S, G, M и выход Типы полового процесса у грибов. Половое раз- множение у грибов известно у всех групп, за исключе- нием анаморфных, или несовершенных, грибов. Поло- вой процесс у грибов разнообразен и его особенности лежат в основе выделения классов. У грибов известно три основных типа полового процесса: гаметогамия, гаметангиогамия и соматогамия (рис. 23). Гаметогамия — слияние гамет, образующихся в га- метангиях. Различают изогамию — слияние подвижных морфологически сходных гамет и гетерогамию — сли- яние подвижных, отличающихся по размерам и часто по степени подвижности гамет. Эти два типа гаметога- мии характерны для хитридиевых и гифохитридиевых грибов. При оогамии крупные, неподвижные яйцеклет- ки, формирующиеся в специальных оогониях, оплодот- воряются мелкими, подвижными сперматозоидами, развивающимися в специализированных антеридиях. У многих грибов с этим типом полового процесса спер- матозоиды не образуются, а яйцеклетка оплодотворяется недифференцированным на сперматозоиды содержимым выростов многоядерного антеридия. Такой тип оогамии характерен для всех представителей отдела оомикота. Второй тип полового процесса, гаметангиогамия, состоит в слиянии двух обычно многоядерных специ- ализированных структур, содержимое которых не диф- ференцировано на гаметы. Гаметангиогамия характер- на для зиго- и аскомикот. Гаметангиогамия у зигомикот носит название зигогамии. Она заключается в слиянии в основном многоядерных клеток, гаметангиев, хорошо отличимых от вегетативного мицелия, на котором они формируются, но недифференцированных морфологи- 1 2 из клеточного цикла состояние Go Показаны соответствующие фазо-стимулирующие факторы (phase-promoting factor), представ- ляющие комлекс циклина (cyclin) — кружок и циклин зависимой протеинкиназы (cydin- dependent kinases — CDK) — квадрат. После прохождения фазы цикла показана деграда- ция циклинов. На рисунке также представ- лен стимулирующий комплекс анафазы (anaphase-promoting complex — АРС), необхо- димый для прохождения одной из стадий ми- тоза — анафазы (расхождение хромосом).
В результате полового процесса образуются гапло- идные, неоднородные в генетическом отношении спо- ры, что принципиально отличает их от спор бесполого размножения грибов. Эти гаплоидные споры распола- гаются или на мицелии или, чаще, на поверхности или внутри плодовых тел различного строения, которые называются телеоморфами. Таким образом, размно- жение с помощью спор, возникших половым путем, даёт начало формам с новой комбинацией генетиче- ского материала, что является основой дальнейшей эволюции форм, а размножение спорами, возникшими бесполым путем, способствует распространению и со- хранению данной формы. чески по половому знаку на мужской и женский. Из об- разовавшейся в результате их слияния зиготы форми- руется одетая толстостенной окрашенной оболочкой зигоспора, которая прорастает после периода покоя в особый зародышевый спорангий (рис. 23, 24). У аскомикот при гаметангиогамии также слива- ются два многоядерных гаметангия, но у них, в отли- чие от зигомикот, половые органы дифференцирова- ны на женский — аскогон и мужской — антеридий. Аскогон состоит из двух клеток: крупной многоядер- ной, или собственно аскогона, и тонкой нитевидной — трихогины, помещающейся на его вершине, через которую в аскогон переливается содержимое много- ядерного антеридия. При этом происходит только плазмогамия, а ядра ассоциируются в пары, образуя дикарион. Из оплодотворенного аскогона без периода покоя вырастают аскогенные дикариотичные гифы. В их клетках происходит слияние ядер дикариона и образование диплоидного ядра, которое в дальней- шем делится мейотически. В результате этого процес- са на аскогенных гифах достаточно сложным путем формируются особые образования — сумки, или аски, внутри которых после митотического деления пост- мейотических ядер формируются восемь эндогенных гаплоидных аскоспор (рис. 25). — — Локализация и морфогенети- ческие изменения в месте обра- зования почки. ФАЗА G1: — Локализация актинового пят- на секреторных элементов в ме- сте возникновения почки. — Начало образования веретена. — Синтез белков РНК, образова- ния кольца и микрофиламентов.
— Рост почки (1/3 размера клет- ки). — Локализация SPA-2, актино- вого пятна и секреторных вези- кул в почке. — SPB веретено в почке.
ФАЗА G2: — Увеличение размера почки до 2/3 размера материнской клетки. — Начало ядерной миграции.
ФАЗА M: — Миграция новых SPB в до- чернюю клетку. — Конденсация хромасом. — Локализация SPA-2 и актина в новом месте. — Образование перегородки. — Цитокинез и отделение клетки.
Рис. 23. Типы полового процесса у грибов [9].
Рис. 24 (1). Жизненный цикл и зигота Rhizopus nigri- cans: а — спорангий, б — спорангиеносец, в — ризоиды, г — зигоспора, д — подвески суспензоры, е — столон.
Третий тип, соматогамия, — половой процесс, при котором сливаются обычные соматические, или вегетативные клетки мицелия. Половые органы и га- меты отсутствуют. Соматогамия характерна для неко- торых представителей отделов хитридиомикота и ги- фохитридиомикота, имеющих одноклеточный таллом. В этом случае целиком сливаются две одноклеточные особи. Такой тип соматогамии называется хологамией. Соматогамия у базидиомикот заключается в слиянии двух вегетативных клеток гаплоидного мицелия. При этом, как и у аскомикот, сначала имеет место только плазмогамия, в результате чего формируются дикари- оны и образуется дикариотический (состоящий из дву- ядерных клеток) мицелий. Это наиболее длительная стадия в цикле развития базидиальных грибов. Затем на этом дикариотическом мицелии формируются осо- бые клетки, базидии, в которых происходит слияние ядер дикариона и мейотическое деление диплоидного ядра, после чего на базидии формируются экзогенные гаплоидные базидиоспоры (рис. 26).
Рис. 24 (2). Жизненный цикл Mucor mucedo. 1 — гапло- идные спорангиоспоры; 2 — вегетативный мицелий; 3 — спорангии, образующиеся на вегетативном однодомным мицелии; 4 — образование зигофор в результате взаимодействия (+) и (-) нитей мицелия; 5 — слияние зигофор; 6 — формирование парагаметангия; 7 — образование ограни- ченного гаметангия с суспензорами; 8 — молодая зигоспора; 9 — зрелая зигоспора; 10 — проростание зигоспоры [11].
Эндогенные аскоспоры сумчатых грибов и экзо- генные базидиоспоры базидиальных грибов образуют- ся в результате полового процесса, т. е. их появление связано с половым размножением этих групп. Половой процесс сумчатых и базидиальных грибов имеет две характерные общие особенности: во-первых, разрыв между плазмогамией и кариогамией и появление ди- кариотической фазы и, во-вторых, отсутствие у зиготы состояния покоя: мейотическое деление диплоидного ядра происходит сразу же после слияния гаплоидных ядер дикариона.
Рис. 25. Половой процесс и развитие сумок у Pyronema omphaloides: 1 — группа половых органов, 2 а — антеридий, б, в — аскогон с трихогиной,
мо комбинирующихся. В случае биполярного опре- деления пола все гифы, выросшие из спор одного плодового тела, распадаются на две группы, и при соединении мицелиев из этих двух разных групп происходит половой процесс. В случае тетрапо- лярного определения пола гифы, выросшие из спор одного плодового тела, распадаются на четыре по- ловые группы. При этом группа I сливается только с группой II, и группа III только с группой IV. Чис- ленные соотношения этих групп для шляпочных грибов (базидиомицетов), у которых тетраполяр- ный гетероталлизм распространён очень широко, соответствует отношению 1:1:1:1 (рис. 27).
По характеру половой дифференцировки у гри- бов различают гомоталличные (обоеполые) и гете- роталличные (раздельнополые) формы. У гомотал- личных грибов к слиянию способны клетки одного и того же мицелия. На одном и том же мицелии формируются и мужской и женский половые ор- ганы (например, оогонии и антеридии у оомикот). У гетероталличных грибов на мицелии, выросшем из одной споры, половые органы не закладываются и, соответственно, зиготы не образуются. Они раз- виваются лишь при встрече двух мицелиев, отлича- ющихся друг от друга по половому знаку (+ и -, или мужской и женский). Понятие гетероталлизма от- носится к гаплоидной стадии, так как определение пола у грибов происходит в основном генотипиче- ски. Гетероталлизм, или раздельнополость, у гри- бов может быть двух типов: биполярный, когда пол определяется одной парой аллелей, и тетраполяр- ный, когда пол определяется двумя парами аллелей, локализованных в разных хромосомах и независи- Рис. 27. Схема распределения пола у грибов.
У несовершенных (анаморфных) грибов половой процесс отсутствует, и жизненный цикл эти грибы проводят в гаплоидном состоянии. В определённой степени отсутствие полового процесса в этой груп- пе компенсируется гетерокариозом и происходящим на его основе парасексуальным процессом. Гетеро- кариоз, или разноядерность, — это наличие в клет- ках мицелия генетически разных ядер. Гетерокариоз характерен для многих групп грибов и обеспечивает адаптацию грибов к изменяющимся условиям среды. В таком мицелии ядра иногда могут сливаться, образуя диплоидное гетерозиготное ядро. Такое ядро делится митотически, при этом происходит митотическая ре- комбинация и затем вегетативная гаплоидизация этих диплоидных ядер путем потери ими части хромосом. Этот сложный процесс, включающий в качестве суще- ственнейшего момента митотическую рекомбинацию, получил название парасексуального процесса. Он известен у разных групп грибов, и имеет особое значе- ние для несовершенных грибов, лишённых настояще- го полового процесса. Особенности деления ядра
Митоз и мейоз у грибов отличаются рядом спец- Функциональная и морфологическая дифференцировка таллома Таблица 5.
ифических особенностей. У большинства видов грибов деление ядра происходит по закрытому типу, то есть Фаза цикла развития Функция Морфологическая структура
Вегетативная Прорастание Проростковые гифы, почкующиеся клетки жгутиковые стадии, у остальных видов веретено де- ления формируется более просто устроенными белко- выми структурами — полярными тельцами веретена (ПТВ). Фазы митоза чередуются быстро, а хромосомы имеют небольшие размеры; в сочетании эти факторы затрудняют микроскопическое исследование, поэтому ранее считалось, что деление ядер у грибов происходит амитотически. Телофаза митоза происходит несинхрон- но, в результате чего могут образовываться гетеропло- идные дочерние ядра, то есть содержащие неравное число хромосом. Чаще всего при гетероплоидии на- блюдается различное число B-хромосом. Митоз и об- разование новых клеток (цитокинез) у мицелиальных (не дрожжевых) грибов происходят независимо друг от друга — ядра перемещаются в дочернюю клетку уже после отделения её перегородкой (септой) от мате- ринской (у грибов с неклеточным мицелием цитокинез вообще наблюдается редко, при регенерации повреж- дённых участков и при образовании репродуктивных органов).
Репродукцион- ная бесполая Разрастание по субстрату Поселение на хозяине
Перенесение неблагоприятных условий Размножение и распространение Столоны, мицелий, ризоморфы, тяжи Апрессории, гифоподии, инфекционные и перфорационные гифы, гаустории, ловчие гифы Хламидоспоры, склероции
Спорангиеносцы (спорангиофоры), спорангиеспоры, конидиеносцы (конидиофоры), конидии, зооспоры
Репродукционное бесполое размножение проис-
|
|||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-05; просмотров: 396; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.20 (0.011 с.) |




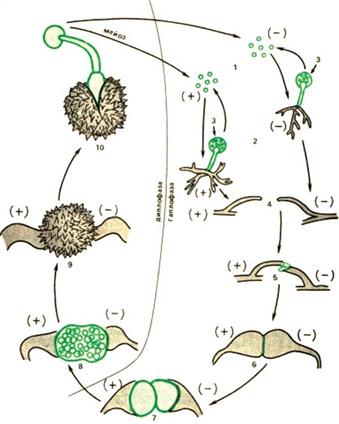
 Начало образования почки. G1
Начало образования почки. G1 ФАЗА S: S
ФАЗА S: S
 G2
G2 M
M




 START
START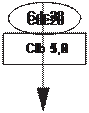


 Рис. 22. Бесполое размножение дрожжей
Рис. 22. Бесполое размножение дрожжей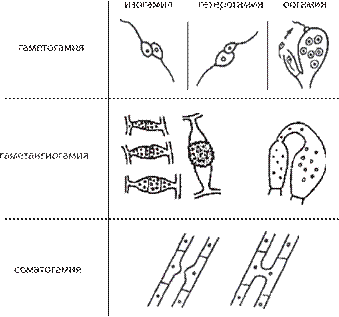



 3 — развитие двуядерных (дикариотичных) аскогенных гиф из оплодотворенного акско- гона, 4 — схема образования сумок из аско- геных гиф с помощью крючка, 5 — молодая и зрелая сумка с аскоспорами [9].
3 — развитие двуядерных (дикариотичных) аскогенных гиф из оплодотворенного акско- гона, 4 — схема образования сумок из аско- геных гиф с помощью крючка, 5 — молодая и зрелая сумка с аскоспорами [9].
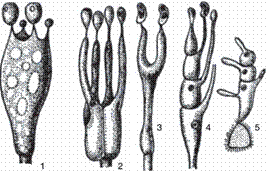

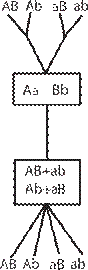


 Рис. 26. Типы базидий: 1 — холобазидия; 2-4 — гетеро- базидии; 5 — телио- (фрагмо-) базидия [9].
Рис. 26. Типы базидий: 1 — холобазидия; 2-4 — гетеро- базидии; 5 — телио- (фрагмо-) базидия [9].
 с сохранением ядерной оболочки. Центриоли имеются лишь у псевдогрибов и некоторых грибов, имеющих
с сохранением ядерной оболочки. Центриоли имеются лишь у псевдогрибов и некоторых грибов, имеющих Гаметы, гаметангии, оогонии, аскогонии, антеридии, трихогины, ооспоры, зигоспоры, аски (сумки), аскоспоры, базидии, базидиоспоры, плодовые тела
Гаметы, гаметангии, оогонии, аскогонии, антеридии, трихогины, ооспоры, зигоспоры, аски (сумки), аскоспоры, базидии, базидиоспоры, плодовые тела


