Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Классификация прокариотических микроорганизмов
В определителе бактерий Д. X. Берджи [3] все про- кариотические микроорганизмы объединены в цар- ство Procaryotae, которое подразделяется на четыре отдела, которые делятся на секции, классы, порядки, семейства, роды, виды. Отдел I. Gracilicutes (от лат. gracilus — тонкий, стройный, cutes — кожа). Включает в себя грамотри- цательные микроорганизмы. В отделе девять секций. С екция 1. Спирохеты. Порядок Spirochaetales. Включает в себя два семейства: Spirochaetaceae (четы- ре рода), Leptospiraceae (один род). С екция 2. Спиралевидные и изогнутые аэро- бы (микроаэрофилы). Одно семейство — Spirillaceae, в котором шесть родов. Патогенны для человека и жи- вотных микроорганизмы рода Campylobacter. С екция 3. Грамотрицательные непод- вижные изогнутые бактерии. Одно семейство — Spirosomonaceae, в котором патогенных три рода. С екция 4. Аэробные грамотрицательные па- лочки, округлые и кокки. Восемь семейств, два из ко- торых имеют патогенные микроорганизмы. Семей- ство Pseudomonadaceae включает в себя четыре рода, более 25 видов, среди которых имеются патогенные (A. mallei и др.). Семейство Neisseriaceae имеет 16 ро- дов. Роды Neisseria и Moraxella содержат патогенные для человека и животных микроорганизмы. Роды Bordetella, Brucella и Francisella не внесены в семейства: содержат патогенные для человека и жи- вотных микроорганизмы. С екция 5. Грамотрицательные факультатив- ные анаэробы. Три семейства: Enterobacteriaceae, Vibrionaceae и Pasteurellaceae. Семейство Enterobacteriaceae имеет 14 родов (Escherichia, Salmonella, Citrobacter, Klebsiella, Enterobacter, Erwinia, Shigella, Proteus, Yersinia и др.). Семей- ство Vibrionaceae имеет два рода. В род Vibrio вклю- чены патогенные микроорганизмы. Семейство Pasteurellaceae имеет три основных рода: Pasteurella, Haemophilus и Actinobacillus. Содержат патогенные виды микроорганизмов. С екция 6. Строгие анаэробы. Изогнутые грамотрицательные палочки. Одно семейство — Bacteroidaceae, в котором 13 родов, среди которых имеются патогенные. С екция 7. Диссимилирующие и разлагающие сульфат бактерии. Семь непатогенных родов. С екция 8. Анаэробные грамотрицательные кок- ки. Одно семейство — Vellonellaceae, в котором три рода. С екция 9. Риккетсии и хламидии. Два поряд- ка: Rickettsiales и Chlamydiales. Порядок Rickettsiales имеет три семейства: Rickettsiaceae, Bartonellaceae и Anaplasmataceae. Семейство Rickettsiaceae имеет три трибы, в которые внесено восемь родов. Семейство Bartonellaceae содержит два рода, a Anaplasmataceae — четыре. Порядок Chlamydiales имеет одно семейство Chlamydiaceae и один род — Chlamydia. Все семейства содержат патогенные микроорганизмы.
Отдел II. Firmicutes (от лат. firmis — крепкий, cutes — кожа). В отдел включены главным образом грамположительные бактерии. С екция 12. Грамположительные кокки. Два семейства: Micrococcaceae и Deinococcaceae. Семей- ство Micrococcaceae имеет четыре рода: Micrococcus, Stomatococcus, Planococcus, Staphylococcus. В секцию кроме указанных двух семейств вне- сены десять самостоятельных родов: Streptococcus, Leuconostos, Pedicoccus, Sarcina и др. С екция 13. Спорообразующие грамположи- тельные палочки и кокки. Шесть родов: Bacillus, Clostridium, Sporolactobacillus, Sporosarcina и др. Пер- вые два рода имеют патогенные виды. С екция 14. Неспорообразующие грамположи- тельные палочки. Семь родов: Lactobacillus, Listeria, Erysipelotrix и др. Имеются патогенные. С екция 15. Неспорообразующие внутри- клеточные грамположительные палочки. 21 род: Corynebacterium, Micobacterium, Propionibacterium, Eubacterium, Asotobacterium, Bifidobacterium, Actinomices и др. С екция 16. Микобактерии. Одно семей- ство Mycobacterioсеае. Семейство имеет один род Mycobacterium, в котором 49 видов: Мус. tuberculosis, Мус. bovis, Мус. avium, Мус. paratuberculosis, Мус. lepra и др. С екция 17. Nocardioforms. Девять родов: Nocardia, Pseudococcus, Pseudonocardia и др. Отдел III. Tenericutes. Объединены грамотрица- тельные прокариоты без клеточной стенки, но име- ющие цитоплазматическую мембрану. В отделе десятая секция — микоплазмы, класса Mollicutes (от лат. molli — мягкий, cutes — покров, кожа). В классе один порядок — Mycoplasmatales — и три семейства: Mycoplasmataceae, Acholeplasmataceae, Spiroplasmataceae. В основном патогенные микоплаз- мы включены в семейство Mycoplasmataceae. С екция 11. Эндосимбионты. Отдел IV. Mendosicutes. Прокариоты, среди ко- торых нет патогенных бактерий; метанобразующие, сероокисляющие, галофилы, микоплазмоподобные, термоацидофильные и другие наиболее древние по происхождению бактерии (архебактерии).
Строение клетки
Клеточная стенка обеспечивает поддержание жесткости структуры клетки, постоянства ее фор- мы и механической прочности. Кроме того, она яв- ляется осмотическим барьером, имеющим зоны из- бирательной проницаемости для веществ различной химической природы. В качестве опорного каркаса она содержит пептидогликан муреин. Основу муреи- на (рис. 6) составляют цепи чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенные β-1,4-гликозидными связями. Остатки мурамовой кислоты соединены полипептидными це- почками, в состав которых входят α-аланин, D-аланин, лизин, D-глутаминовая и мезодиаминопимелиновая кислоты.
Аминокислоты D-ряда и мурамовая кислота уни- кальны для прокариот, своеобразие структуры клеточ- ной стенки служит основой избирательного действия некоторых антибиотиков, например пенициллина и других β-лактамов. Клеточные стенки грамположи- тельных и грамотрицательных бактерий существенно различаются по своей структуре. А В
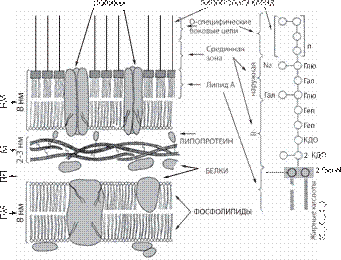
D-ala D-ala D-ala Рис. 6. Пептидогликан E. coli: A-N-ацетилмурамовая кислота; B-N-ацетил- глюкозамин; ala — ала- нин; glu — глутаминовая кислота; DAP — диа- минопимелиновая кислота. У грамотрицательных бактерий муреиновая сеть однослойная, иногда двуслойная и составляет не бо- лее 10% сухой массы клеточной стенки. На ней рас- полагаются белки, липопротеиды, липополисахариды и фосфолипиды, входящие в состав внешней мем- браны (рис. 7). Стабилизация этих компонентов обе- спечивается ионами Са2+ и Mg2+. Существенное зна- чение для структуры и функции внешней мембраны имеет липид А. Его скелет содержит дисахарид, со- стоящий из остатков D-глюкозамина, соединенных β -1,6-связью, имеющих в положении 1 и 4 фосфатные группы. Скелет этерифицирован жирными кисло- тами С12, С14 и С16. Липид А имеет уникальную кон- формацию — компактную и высокоупорядоченную, благодаря чему создает в мембране вязкую структуру, которая затрудняет диффузию желчных кислот, детер- гентов и некоторых антибиотиков. Липид А обеспечи- вает токсичность и пирогенность липополисахарида. Антигенная специфичность грамотрицательных бак- терий, главным образом, определяется углеводами О-замещенных боковых цепей, выступающих наружу с поверхности клетки. Внешняя мембрана определяет высокую устойчивость грамотрицательных бактерий по сравнению с грамположительными к антимикроб- ным агентам.
Рис. 7. Модель строения цитоплазматической мем- браны грамотрицательных бактерий: Справа представлена липополисахаридная моле- кула. Глю — глюкоза; Глю-N — глюкозамин; NA = N-АцГлю-N-ацетилглюкозамин; Гал — галактоза; Геп — гептоза; КДО — 2-кето- З-дезоксиоктоновая кислота; М — муреин; НМ — наружная мембрана; ПМ — плазмати- ческая мембрана; ПП — периплазматическое пространство [7].
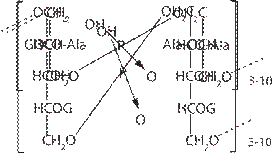
Антибиотики широкого спектра, например бета- лактамы, вызывают освобождение липополисахари- дов из внешней мембраны, что может привести к эн- дотоксическому шоку у больного. У грамположительных бактерий внешняя мембра- на отсутствует, а муреиновая сеть составляет 30-70% сухой массы клеточной стенки и достигает 40 слоев. Характерно наличие тейхоевых и тейхуроновых кис- лот (рис. 8), обеспечивающих отрицательный заряд клетки и способствующих сорбции катионов из окру- жающей среды. У некоторых микроорганизмов могут присутствовать добавочные компоненты — липиды, воска, миколовые кислоты, протеины, полисахариды.
Рис. 8. Модель строения цитоплазматической мем- браны Гр+ бактерий. А — глицеролтейхоевая кислота; В — рибитолтейхоевая кислота G — гликозил, Ala — аланил.
Различие в структуре клеточной стенки двух групп микроорганизмов выявляют с помощью окрашивания по Граму. Препарат обрабатывают раствором кристал- лического фиолетового, затем йода. Образующийся комплекс красителя с йодом располагается на протопла- сте. При обработке препарата спиртом он удерживается клеточной стенкой грамположительных бактерий и вы- мывается — у грамотрицательных. Способность окра- шиваться по Граму — важный таксономический при- знак, с которым коррелируют другие свойства бактерий. S-слой (surface — поверхность) располагается на поверхности клеток всех прокариот и покрывает целиком всю клетку. Он состоит из структурных еди- ниц — протеинов или гликопротеинов, образующих монослой, структура которого типична для двухмер- ных кристаллов (решетка гексагональной, косой или квадратной симметрии). Взаимодействие между субъ- единицами и подлежащими структурами происходит за счет нековалентных связей. S-слой обеспечивает за- щиту клетки от внешних воздействий, однако при про- должительном культивировании он может быть утра- чен без потери жизнеспособности штамма. Капсулы и слизь образуются у некоторых бактерий снаружи от клеточной стенки, как ее внешний слой. Способность к их формированию не является видо- вым признаком: могут существовать капсульные и бес- капсульные штаммы. У патогенных микроорганизмов капсула обеспечивает защиту от фагоцитоза, повышая вирулентность штамма (у пневмококков). У микробов, обитающих в почве и на растениях, капсула защищает клетки от высыхания, солнечной радиации, биоцидов. Капсулы и слизь создают для микробных клеток ос- мотические условия, благоприятствующие сорбции питательных веществ из субстрата, способствуют ад- гезии клеток между собой и субстратом. Многие эк- зоферменты локализуются в капсуле, где происходят превращения веществ, поступающих в клетку (рис. 9).
Рис. 9. Капсулы вокруг клеток Clostridium.
У большинства бактерий капсулы и слизь имеют полисахаридную природу. У некоторых бацилл это по- липептиды в основном D- и L-глутаминовой кислоты. Капсульные полисахариды обладают антигенной специфичностью и используются для изготовления вакцин (у пневмококков, менингококков), для иден- тификации и классификации (у сальмонелл). Рас- творимые слизи (декстран Leuconostoc dextranicum, L. mesenteroides, ксантан Xanthomonas campestris) получают в промышленных масштабах и широко ис- пользуют в фармации и других областях.
Протопласты и сферопласты — это структуры, полностью (протопласты) или частично (сферопла- сты) утратившие клеточную стенку, например, под действием лизоцима или пенициллина. Это осмотиче- ски лабильные элементы, которые могут существовать только в гипертонических растворах. Они сохраняют биологическую активность и способны в специаль- ных условиях ревертировать в нормальные клетки. Используются в клеточной инженерии для получения гибридных форм микроорганизмов. L-формы, получившие свое название в честь инсти- тута Листера в Лондоне, образуются в условиях, приво- дящих к нарушению синтеза клеточной стенки; напри- мер, у больного туберкулезом возбудитель под влиянием лекарственных веществ может превратиться в L-форму. При этом микобактерии теряют характерную кислото- устойчивость, что затрудняет их выявление и диагно- стику заболевания. Для таких клеток характерны не- правильные формы, иногда нитевидные, способные проходить через поры бактериальных фильтров. Лабиль- ные L-формы способны ревертировать в нормальные клетки. Стабильные формы не образуют клеточной стен- ки, поскольку ее утрата связана с изменением генотипа (мутацией). Периплазматическое пространство располагается между слоем муреина и цитоплазматической мембра- ной. В нем находятся ферменты гидролазы, расщепля- ющие вещества, поступающие в клетку, и полимеразы, участвующие в синтезе клеточной стенки и капсулы, а также белки, принимающие участие в транспорте субстратов в цитоплазму, и белки — рецепторы хемо- таксических стимулов. Цитоплазматическая мембрана располагается под клеточной стенкой и отделяет от внешней среды ци- топлазму. Имеет толщину 6-8 нм и составляет 8-15% сухого вещества клетки. Ее структура соответствует общему принципу организации мембран про- и эука- риотических клеток. Она состоит из двух слоев моле- кул липидов, у которых гидрофобные цепочки жирных кислот ориентированы перпендикулярно ее плоскости, а гидрофильные полярные части молекул соединены с молекулами белков за счет полярного, электростати- ческого и гидрофобного взаимодействия. Молекулы липидов и белков не сохраняют фиксированной ори- ентации, но находятся в постоянном движении внутри остова данной мембраны (рис. 10) Структура грамположительной клеточной стенки. Клеточная стенка содержит до 40 слоев пептидогли- кана. Молекулы тейхоевых кислот ковалентно связа- ны с пептидогликаном (ПГ). Липотейхоевые кислоты (ЛТК) содержат липидные «хвостики», закрепленные в гидрофобной области ЦПМ. Клеточные стенки могут иметь белковые слои на поверхности. Белковые струк- туры располагаются или островками (как показано на схеме), или тесно упаковываются, образуя S-слой. Структура грамотрицательной клеточной стенки. Тонкий слой пептидогликана сверху покрыт внешней мембраной (ВМ), прикрепленной к нему липопроте- идами (ЛП). Между двумя мембранами — периплаз- матическое пространство со слоем пептидогликана (ПГ) внутри него. Внешняя мембрана на внутренней поверхности содержит фосфолипиды, на внешней — липополисахариды (ЛПС). ЛПС состоят из липидной части, обращенной внутрь внешней мембраны, фор- мируя ее гидрофобную область, и полисахаридной ча- сти, обращенной во внешнюю среду.
Липиды, в основном, полярные фосфолипиды, со- ставляют около 40% массы мембраны. Они выполня- ют структурные функции и обеспечивают конформа- ционные изменения молекул ферментов, необходимые для проявления их активности.
Рис. 10. Структура бактериальной клеточной стенки. Вверху: грамположительная клеточная стен- ка. 1. Цитоплазматическая мембрана, 2 — ПГ, 3 — фосфолипид, 4 — протеин, 5 — липотей- хоевая кислота. Внизу: грамотрицательная клеточная стенка. 1 — внутренняя мембрана, 2 — периплазматическое пространство, 3 — внешняя мембрана, 4 — фосфолипид, 5 — ПГ, 6 — ЛП, 7 — протеин, 8 — ЛПС, 9 — порины.
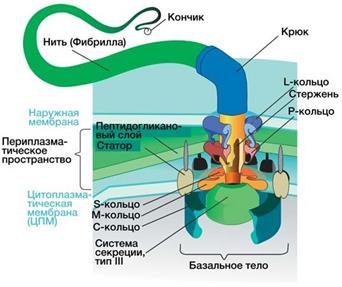
Белки мембран подразделяют на периферические и интегральные. Первые находятся на поверхности, вторые погружены в толщу мембраны. Белки-порины выстилают поры мембран, обеспечивая ее проницае- мость. Определенные белки мембран выполняют ката- литические функции или являются рецепторами, свя- зывающими вещества, необходимые для клетки. Цитоплазматическая мембрана служит осмотиче- ским барьером, в ней локализуются системы активно- го транспорта веществ в клетку и из клетки. Мезосомы — особые структуры, образуемые путем инвагинации (впячивания) мембраны, содер- жат ферменты системы окислительного фосфорили- рования и выполняют у прокариот функции мито- хондрий. Цитоплазма составляет внутреннюю среду клетки. Это сложная высокогетерогенная система, в ней распо- лагается генетический материал клетки (нуклеоид, плаз- миды), 70 S рибосомы, ферментные системы, выполня- ющие метаболические функции, резервные вещества (полисахариды, липиды, полигидроксимасляная кисло- та, полифосфаты, сера у серобактерий). Основная часть метаболических процессов осуществляется в цитоплаз- ме. Мембранные структуры, располагающиеся в цито- плазме, наиболее развиты в клетках эукариот, у бактерий имеются их аналоги (мезосомы, вакуоли, лизосомы). Жгутики предназначены для передвижения бак- терии (рис. 11). Перемещаться без жгутиков способны цианобак- терии, скользящие бактерии и спирохеты. Число жгу- тиков и их расположение на клетке — таксономиче- ский признак, характерный для определенных видов. Монотрихи — бактерии с одним жгутиком на кон- це (Vibrio cholerae); амфитрихи имеют полярно распо- ложенные жгутики на двух концах (Spirillum volutans); лофотрихи — пучок жгутиков на одном конце (Alcaligenes faecalis); у перитрихов жгутики располо- жены по всей поверхности клетки (E. coli, Salmonella spp.) (рис. 12).
Б
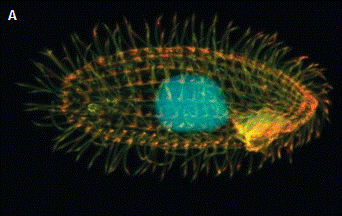
Рис. 11. Органоиды движения у микроорганизмов: А — Tetrahymena thermophila; Б — строение жгути- ка бактерий.
Жгутики построены из белка — флагеллина, их диаметр 10-20 нм, длина до 20 мкм. Жгутик закре- плен в цитоплазматической мембране и клеточной стенке с помощью базального тельца, состоящего из центрального стержня и двух пар (у грамотрица- тельных бактерий) и одной пары (у грамположитель- ных) дисков. Жгутики вращаются благодаря тому, что через диски проходит поток заряженных частиц (Н+, ОН–, Na+) за счет разности потенциалов внутри и вне клетки. Жгутики находятся под контролем си- стемы, воспринимающей информацию о состоянии окружающей среды. Поэтому они позволяют клет- кам перемещаться в область с оптимальными ус- ловиями (таксис). Существует около 35 генов, уча- ствующих в сборке и функционировании базального тельца и еще более 20 генов, определяющих направ- ление движения в ответ на внешние стимулы. Вся система находится под контролем главного оперона, который регулируется системой цАМФ — белок-ак- тиватор.
Рис. 12. Основные типы расположения жгутиков бактерий [4]
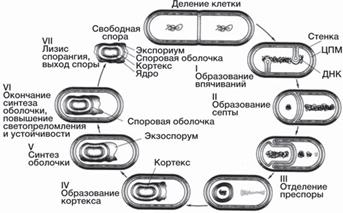
Фимбрии (пили или волоски) располагаются на поверхности клеток многих бактерий. Их чис- ло на клетке может доходить до 10 000. Их диаметр 3-25 нм, длина около 12 мкм. Они обеспечивают сце- пление клеток, например при конъюгации, их адгезию к субстрату. Белки фимбрий служат рецепторами, обе- спечивающими биологическое узнавание. Некоторые фимбрии являются капиллярами, связанными с мезо- сомами, и участвуют в водно-солевом обмене. F-волоски (F-пили) по структуре напоминают фимбрии, однако на клетке их не более одного-двух. Они находятся лишь у клеток, способных к передаче генетического материала при конъюгации, и принима- ют участие в этом процессе как рецепторы и связыва- ющие структуры. Кроме того, они являются рецепто- рами специфических фагов. Споры прокариот — это особая структура, пред- назначенная для сохранения в неблагоприятных усло- виях. Споры по сравнению с вегетативными клетками намного устойчивее к воздействию высокой темпера- туры, радиации, химических агентов. Споры образу- ются внутри бактериальной клетки обычно при исто- щении в культуральной среде питательных веществ и накоплении продуктов обмена. Биохимическим сигналом для спорообразования служит снижение концентрации в клетке гуаниловых нуклеотидов — ГТФ и ГДФ. Спорообразование зависит от плотности популяции: при малой концентрации клеток споры не образуются. Спорообразование восстанавливает- ся, если к такой культуре добавить фильтрат поздней экспоненциальной фазы культуры, в которой клетки начали спорулировать (см. чувство кворума). Обра- зование спор требует синтеза ДНК: блокада реплика- ции прекращает экспрессию ранее индуцированных генов споруляции. Образование споры начинается с накопления белкового материала, при этом расходу- ются запасные питательные вещества. Белки споры содержат значительно больше цистина, чем вегета- тивные клетки. Предполагают, что многочисленные дисульфидные связи в белке обеспечивают высокую механическую прочность оболочек спор. На ранней стадии споруляции образуются особые белки, кото- рые связываются с ДНК, слегка раскручивая ее, что изменяет геометрию пиримидиновых оснований и повышает их устойчивость к ультрафиолетовому излучению. В период споруляции образуется спец- ифическое вещество — дипиколиновая (пиридин- 2,6-дикарбоновая) кислота, которая в виде соли каль- ция входит в состав оболочки споры. Морфологические изменения клетки в процессе споруляции (рис. 13) начинаются с образования асим- метрично расположенной септы. Подобно обычной септе, образующейся при делении клеток, она состоит из двух мембран с тонким слоем пептидогликана меж- ду ними. Большая часть клетки является материнской, а меньшая — проспорой, содержащей нуклеоид. Далее септа деградирует, а мембрана материнской клетки об- волакивает проспору, которая таким образом получает двойную мембрану. Между этими мембранами распо- лагается слой пептидогликана, формирующий кортекс споры. На следующей стадии спорообразования про- исходит сборка белковых элементов оболочки споры. По окончании созревания споры материнская клетка лизируется. Зрелая спора содержит минимальное ко- личество свободной воды и повышенное по сравнению с вегетативной клеткой количество липидов. На долю ее оболочки приходится до 50% сухой массы. Все эти особенности обеспечивают ее устойчивость к факто- рам внешней среды. Спорообразование присуще преимущественно па- лочковидным микроорганизмам (бациллам, клостри- диям). К ним относятся возбудители сибирской язвы, столбняка, анаэробной инфекции, ботулизма и неко- торые сапротрофные виды. Помимо этого споры об- разуют виды родов Thermoactinomyces, Sporosarcina, Sporomusa. Все они за исключением последнего по Граму окрашиваются положительно. Спора — это покоящаяся форма. В благоприятных условиях споры прорастают. При этом спора набухает, поглощая воду, возрастает ее метаболическая актив- ность, выделяется дипиколиновая кислота, спора утра- чивает свою устойчивость. Наконец, оболочка споры разрывается, и из нее выходит вегетативная клетка (рис. 13).
Рис. 13. Цикл развития Bacillus subtilis: 1 — вегетатив- ная клетка; 2 — образование септы споруля- ции; 3-4 — образование двойной мембраны споры; 5 — формирование кортекса (а); 6 — формирование белковой оболочки (б); 7 — зрелая спора; 8 — прорастание споры [2].
Способность микроорганизмов к спорообразо- ванию учитывают при выборе методов дезинфекции и стерилизации, имея в виду высокую устойчивость спор к биоцидным агентам. Наиболее устойчивые виды используют в качестве тест-культур для оценки эффек- тивности стерилизации: Bac. stearothermophilus — па- ром под давлением, некоторые виды Вас. subtilis — су- хим паром, Bac. pumilus — радиационной. Клетки прокариот (от греч. pro — до, karion — ядро) не имеют оформленного ядра. Иными словами, генетический материал (ДНК) прокариот находится прямо в цитоплазме и не окружен ядерной мембраной.
1.6 Бактериальная хромосома представляет собой замкнутую кольцевую ДНК длиной около 1,3 м. ДНК бактерий представлена одиночными кольце- выми молекулами длиной около 1 мм. Каждая такая молекула состоит примерно из 5´106 пар нуклеотидов. Суммарное содержание ДНК (геном) в бактериальной клетке намного меньше, чем в эукариотической, а сле- довательно, меньше и объем закодированной в ней ин- формации. В среднем, такая ДНК содержит несколь- ко тысяч генов, что примерно в 500 раз меньше, чем в клетке человека. Плазмиды и эписомы Плазмиды и эписомы — это небольшие фрагменты ДНК, отличающейся от основной массы ДНК. Они ча- сто реплицируются вместе с ДНК хозяина, но не нуж- ны для выживания его клетки. Сначала было принято различать эписомы и плаз- миды: эписомы внедряются в ДНК хозяина, а плазми- ды — нет. К эписомам относятся F-факторы и так назы- ваемые умеренные фаги. Сейчас обе группы называют одним общим термином «плазмиды». Плазмиды ши- роко распространены в природе, и в последние годы их считают внутриклеточными паразитами или симби- онтами, устроенными еще проще, чем вирусы. Вопрос о том, можно ли вирусы считать живыми организма- ми, будет подробно рассмотрен далее. Что касается плазмид, то здесь дело обстоит еще сложнее — ведь они представляют собой только молекулы ДНК. Плаз- миды контролируют свою репликацию и число копий в клетке, которое у разных плазмид может колебаться от 1 до 100. Для плазмид характерно явление несо- вместимости, т. е. неспособности близкородственных плазмид существовать в одной клетке; на этом осно- вана их классификация. Основная таксономическая единица у плазмид — Inc-группа (incompatibility — несовместимость). Плазмиды, относящиеся к одной 1 пс-группе, обладают многими общими свойствами: сходной молекулярной массой, высокой степенью го- мологии ДНК — и контролируют сходные фенотипи- ческие признаки. Механизм несовместимости состоит в угнетении репликации проникшей в клетку плазмиды специфиче- ским белком или РНК, вырабатываемым под контролем плазмиды, имеющейся в клетке. Кроме того, под кон- тролем последней на поверхности клетки синтезирует- ся специфический белок или липопротеин, препятству- ющий вхождению в клетку родственной плазмиды. Плазмиды придают своим клеткам-хозяевам це- лый ряд особых свойств. Некоторые плазмиды явля- ются «факторами резистенции» (R -плазмиды, или R -факторы: от англ. R = resistance — устойчивость), т. е. факторами, придающими устойчивость к антибио- тикам. Примером может служить пенициллиназная плазмида стафилококков, которая трансдуцируется различными бактериофагами. В этой плазмиде содер- жится ген, кодирующий фермент пенициллиназу, кото- рая разрушает пенициллин и, таким образом, придает устойчивость к пенициллину. Передача и распростра- нение таких факторов среди бактерий (в результате полового размножения) очень мешают врачам. Другие плазмидные гены определяют устойчивость к дезин- фицирующим средствам, способствуют таким забо- леваниям, как стафилококковая импетиго; помогают молочнокислым бактериям превращать молоко в сыр; придают способность усваивать такие сложные веще- ства, как углеводороды, что можно использовать для борьбы с загрязнениями океана или для получения кормового белка из нефти.
|
||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-05; просмотров: 157; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.238.70 (0.063 с.) |