Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Взаимодействие вируса с клеткой хозяина
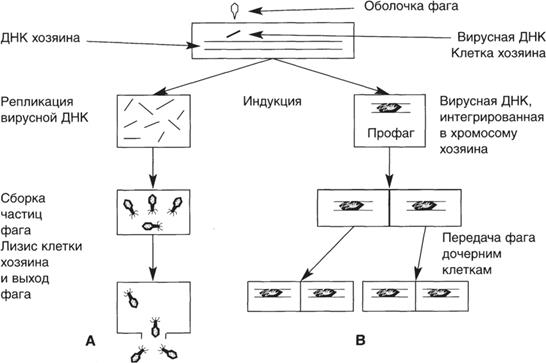
Вирус проникает в клетку хозяина и использует ее метаболические системы для своего воспроизвод- ства и сохранения [4]. Этапы этих процессов различ- ны у разных групп вирусов и окончательный результат может быть одним из следующих: 1. Размножение вируса и гибель клетки хозяина. 2. Размножение вируса и выход его из клетки без значительного повреждения последней. 3. Сохранение вируса в клетке в латентном состо- янии, обычно в виде вирусной НК. 4. Внедрение вирусной НК в геном хозяина, в ре- зультате чего происходит мутация, например, возник- новение раковых клеток. Бактериофаги (фаги) — это вирусы бактерий (прокариот). Их генетический материал содержит- ся в головке, имеющей белковую оболочку (рис. 37). Хвостовые нити и зубцы предназначены для распоз- навания рецепторов на поверхности бактериальной клетки и адсорбции на ней. Распознавание специфич- но не только для вида бактерии, но и для штамма, что служит основой для фаготипирования бактерий (см. ниже). У некоторых фагов хвост имеет чехол, по- крывающий стержень. После адсорбции фага на клет- ке чехол сокращается, проталкивая стержень внутрь. Через стержень фаговая НК проникает внутрь клет- ки. Процесс облегчается благодаря местному по- вреждению клеточной стенки фаговым лизоцимом. Таким сложным органом инфицирования обладают лишь некоторые фаги грамотрицательных бактерий. На клеточной стенке грамположительных бактерий имеются рецепторные участки, которые способствуют проникновению в клетку крупных молекул и бактери- офагов. Чувствительное место для атаки — это пили, к которым фаги могут прикрепляться. Некоторые фаги впрыскивают в клетку свою НК, другие проникают ин- тактными. По типу взаимодействия с бактериальной клеткой фаги подразделяют на вирулентные и уме- ренные. Вирулентные фаги размножаются внутри клетки. Созревшие частицы фага изнутри разрушают клеточную стенку и выходят наружу, клетка при этом погибает. Умеренные фаги также способны лизировать бактерии, однако, в большинстве клеток популяции они существуют в клетке в виде профага — фаговой НК, которая подобно плазмидам может интегрировать с хромосомой. Литический цикл жизни вирулентного ДНК- геномного фага начинается с проникновения его ДНК в клетку и синтеза ранней иРНК (ранними называют- ся молекулы, образующиеся до репликации вирус- ной НК). На матрице вирусной иРНК синтезируются ранние белки. Последние выключают синтез белков клетки хозяина, разрушают бактериальную ДНК и на- чинают синтез компонентов вирусной ДНК. После
этого происходит репликация вирусной ДНК, синтез поздней иРНК и белков, необходимых для построения фаговых частиц (капсомеров головки и элементов хво- ста) и лизоцима. Вновь синтезированные структурные единицы путем самосборки образуют зрелые фаговые частицы, покидающие клетку, оболочка которой раз- рушена лизоцимом. В одной микробной клетке мо- жет быть синтезировано до 100 фаговых частиц через 25 мин после инфекции. Литическую активность вирулентного фага можно выявить в эксперименте, путем посева смеси суспен- зии фага и чувствительной к нему культуры на пита- тельный агар в чашке Петри. На газоне бактериальной культуры появятся зоны лизиса в результате гибели части клеток под действием фага. Поскольку каждая зона лизиса (колония фага) инициирована одной фаго- вой частицей, метод позволяет определить их количе- ство в исходной суспензии (метод титрования фага). Лизогенными называют культуры, несущие уме- ренный фаг. Чтобы обнаружить явление лизогении, т. е. выход фага из клеток, требуется индикаторная культу- ра, для которой данный фаг вирулентен. Лизогенную культуру смешивают с избытком бактерии-индикатора и высевают газоном на чашку. Наблюдают зоны лизиса индикаторной культуры под действием фага. В центре такой зоны находятся клетки лизогенной культуры. Основные стадии развития умеренных и вирулентных фагов представлены на рис. 38. Интеграция фаговой НК с бактериальной хромо- сомой обеспечивает ее передачу дочерним клеткам. Лизогенные бактерии невосприимчивы к заражению теми фагами, которыми они лизогенизированы, а так- же близкородственными фагами. Эта невосприимчи- вость связана с образованием особого репрессора, пре- пятствующего размножению фага. Этот же репрессор препятствует переходу профага в активное состояние и синтезу фаговых белков. Спонтанно лизогенные бактерии лизируются редко (10–2-10–5 в одной генерации). Частота лизиса зависит от внешних условий, например, состава пи- тательной среды. Мутагены (ультрафиолетовые лучи, Н2 О2, митомицин С и др.) могут индуцировать массо- вое развитие зрелых фаговых частиц в клетках лизо- генной культуры, связанное с нарушением механизма репрессии. Мутации также могут быть причиной пере- хода умеренного фага в вирулентное состояние. Такие мутанты оказываются устойчивыми к репрессору или утрачивают способность вызывать синтез репрессора в клетке.
Обычно лизогения — это весьма стабильное со- стояние, однако, некоторые клетки способны утрачи- вать фаг и вместе с этим резистентность к данному типу фага. Лизогения — чрезвычайно распространенное яв- ление: большая часть штаммов бактерий несет в себе НК одного или нескольких фагов, которая определяет
Рис. 38. Схема развития вирулентного (А) и умеренного (В) бактериофагов. фенотипические показатели культуры (морфологиче- ские, культуральные, антигенные, токсигенные и др.). Это явление носит название фаговой конверсии. Инфекционные фаги, продуцируемые лизогенной культурой, способны лизогенизировать другие штам- мы данного вида бактерии (или близкородственных видов). При переходе из интегрированного с бакте- риальной хромосомой состояния в автономное ге- ном фага может включить в свою структуру соседние гены нуклеоида клетки донора и перенести их в дру- гую клетку (реципиент). Это явление носит название трансдукции. Путем трансдукции могут быть переда- ны многие важные признаки бактерий: резистентность к антибиотикам, вирулентность, токсигенность и др. Практическое использование фагов. Фаги широ- ко используются в генетической инженерии в качестве векторов — переносчиков генов в процессе создания рекомбинантных молекул ДНК. В медицине фаги на- значают с профилактической и лечебной целью при дизентерии, брюшном тифе и других энтеральных за- болеваниях, при гнойно-воспалительных процессах и дисбактериозе. Широко используют фаги в диагно- стике инфекционных заболеваний и идентификации микроорганизмов. Реакция нарастания титра специ- фичного фага указывает на присутствие соответствую- щего вида микроорганизма в объектах внешней среды (вода, пищевые продукты и т. п.). Метод фаготипиро- вания позволяет установить биовар бактерии и тем са- мым выявить источник инфекции. Поскольку многие вещества, вызывающие индукцию профага и переход его в активное состояние, являются онкогенными, ли- зогенные культуры бактерий могут быть использова- ны для выявления потенциальных канцерогенов. Размножение вирусов млекопитающих. По сравнению с бактериофагом, литический цикл которого завершается в пределах 30 мин, виру- сы млекопитающих размножаются медленно, в куль- туре ткани цикл репликации занимает от 4 до 24 час и включает стадии адсорбции, проникновения внутрь клетки и процесс образования зрелых вирусных ча- стиц. Адсорбция обусловлена двумя механизмами: не- специфическими (электростатическими и ван-дер- ваальсовыми силами) и специфическими, более прочными, представляющими собой взаимодействие рецепторов вируса с соответствующими рецепторами клетки по принципу биологического узнавания. Проникновение вирусов млекопитающих внутрь клетки зависит от природы вируса. На поверхности вирионов многих групп вирусов, например, гриппа имеются особые шипы, содержащие нейраминидазу и гемагглютинин, которые участвуют в проникнове- нии вириона в клетку. Вирусы оспы и герпеса погло- щаются клеткой, как при фагоцитозе.
Депротеинизация (высвобождение вирусной НК) происходит с участием ферментов клетки хозяина. Синтез вирусных НК и белков определяется при- родой вируса. У ДНК-геномных вирусов процесс начинается с синтеза ранней иРНК с участием РНК- полимеразы клетки хозяина или вириона. На матрице ранней РНК синтезируются ранние белки, необходи- мые для последующей репликации ДНК. Реплика- ция также происходит под действием клеточных или вирусных ферментов. На матрице реплицирующейся ДНК происходит синтез поздних иРНК, которые на- правляют синтез белков вируса. У РНК-геномных вирусов, содержащих +РНК, по- следняя транслируется на рибосомах клетки хозяина. Вирусная –РНК используется как матрица для постро- ения с помощью РНК-зависимой РНК-полимеразы комплиментарной копии +РНК, которая функциониру- ет как информационная. Необходимым этапом жизненного цикла ретро- вирусов является интеграция его генома в форме ДНК-провируса в хромосому хозяина. Синтез ДНК- провируса на матрице вирусной +РНК происходит с участием РНК-зависимой ДНК-полимеразы (обрат- ной транскриптазы). Интегрированная в одну из хро- мосом хозяина вирусная ДНК транскрибируется клеточной РНК-полимеразой. Ретровирусы часто яв- ляются онкогенными, поскольку включение их ДНК в геном клетки-хозяина вызывает ее перерождение. По этой же причине онкогенными могут быть и ДНК- геномные вирусы. Самосборка вириона — это физико-химический процесс, в результате которого формируется капсид с встроенной в него НК. У вирусов, имеющих наруж- ную оболочку, формирование вирионов происходит на клеточной мембране, компоненты которой входят в состав оболочки вируса (рис. 39).
Рис. 39. Схема сборки и выхода дочерних популяций вируса гриппа из зараженных клеток [24].
Выход вирионов у одних вирусов сопровождается гибелью клетки, у других — только частичным по- вреждением мембраны. Культивирование вирусов
Культивирование вирусов в лабораторных услови- ях является необходимым этапом диагностики многих вирусных болезней, кроме того, оно необходимо для получения вакцинных препаратов. Поскольку виру- сы являются облигатными паразитамми, они способ- ны размножаться только в живых клетках, например, в культуре ткани (клетках тканей человека или жи- вотных, растущих на питательном субстрате, обыч- но в виде монослоя на плоской поверхности сосуда). Присутствие вирусов можно обнаружить по цитопа- тическому эффекту (ЦПЭ), т. е. разрушению монослоя клеток. Метод позволяет идентифицировать вирус, например, в клиническом материале, с помощью им- мунной сыворотки. Специфическая сыворотка нейтра- лизует вирус, и ЦПЭ в ее присутствии не будет про- являться. Вирусы выращивают также путем заражения ла- бораторных животных или эмбрионов птиц.
|
||||||||||
|
|
Последнее изменение этой страницы: 2021-04-05; просмотров: 155; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.116.159 (0.012 с.) |