Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Цветовая чувствительность (или цветовое зрение)Содержание книги
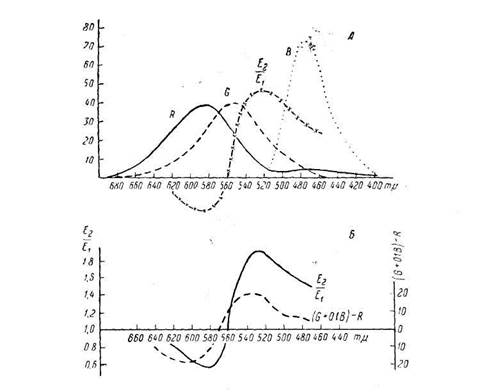
Поиск на нашем сайте Особенно много времени и внимания уделил С. В. Кравков проблеме цветового зрения. В предисловии к книге «Цветовое зрение» С. В. Кравков формулирует свою задачу как стремление выяснить то место и положение, которое занимает вопрос о цветовом зрении в системе марксистско-ленинской гносеологии. С другой стороны, автор хотел показать, что дальнейший прогресс науки о цветовом зрении представляет собой в сущности уточнение и развитие трехкомпонентной теории цветового зрения, впервые сформулированной Ломоносовым. В книге конкретизируются, применительно к цветовому зрению, идеи ионной теории возбуждения, развивавшейся в свое время П. П. Лазаревым, и излагаются новые и нигде ранее не описанные данные о действии побочных раздражителей на цветовое зрение. Таким новым фактом является противоположная реакция цветоощущающего аппарата глаза человека на ряд раздражителей, а именно: адреналин, звук, электротон, запахи. Работы эти были начаты в 1936 г. 1 G. Studnitz, Physiologie des Sehens, 1952. J. London. Reaserch on sensory interaction in the Soviet Union Psycholog. Bull., v. 51, № 6, 531 -586, 1954. A. D u Bois P о u 1 si n. Le Champ visuel, 1953. Y. Segal. Mechanismus der Farben Sehens. 1957. Ive Le Grand. Optigue Physiologie, 1948. 90 Измеряя цветовую чувствительность на монохроматоре для темно-адаптированного глаза, Кравков нашел, что под влиянием адреналина, звука и анэлектротюна повышается чувствительность к излучениям коротковолновой части спектра и понижается чувствительность к длинноволновой части спектра, т. е. повышается чувствительность к зелено-синим лучам и понижается к оранжево-красным. То же наблюдалось при измерении критической частоты слития мельканий для спектральных цветов. При обонятельных раздражениях критическая частота слития мельканий для зеленых лучей снижалась, а для красных повышалась. Добрякова [31] в результате применения вкусовых (сладкое) и температурных раздражений (тепла на руку) получила такие же взаимнопротивоположные изменения в коротко- и длинноволновой части спектра критической частоты слития мельканий для слабых яркостей, при которых она (рав'няш'ась 12 – 18 кол/сек, для больших же яркостей, порядка 26 – 30 кол/сек, наблюдалась обратная картина – повышение критической частоты для зеленых и понижение для красных лучей. Исследуя одновременно в одном опыте цветовую чувствительность и критическую частоту, Кравков нашел, что повышению чувствительности к зеленому цвету соответствует снижение критической частоты слития мельканий. Следовательно, описанное выше снижение критической частоты в зеленых лучах от запахов, тепла и вкусовых раздражителей нужно понимать как повышение чувствительности к зеленым лучам, так же как от звука, анэлектротона и адреналина. В работах Л. И. Селецкой [77] было показано, что под влиянием размарина и индола противоположно меняются поля зрения для красного и зеленого цвета. Под влиянием размарина граница поля зрения для зеленого суживается, а для оранжево-красного расширяется. Под влиянием индола, наоборот, суживаются поля для красного, а для зеленого расширяются. При запрокинутой голове чувствительность к зеленому снижается, чувствительность к красному – повышается [107]. В чем же здесь дело? Оказалось, что первая группа раздражителей – слуховые, запах гераниола и др. – возбуждает по преимуществу симпатическую нервную систему, подобно адреналину, кардиамину и т. п. (Кравков, Шварц). Вторая же группа раздражителей – запах индола, запрокидывание головы – возбуждает по преимуществу парасимпатическую нервную систему, подобно пилокарпину, карбохолину, китайскому лимоннику (Кравков, Селецкая, Галочкина, Шварц). Таким образом, все вышеописанные работы говорят о противоположной вегетативной природе красно- и зеленоощущающего аппаратов глаза. Это подтвердили и опыты Кравкова и Галочкиной [55], [56] с действием ан- и катэлектротона и ионофореза кальция и калия на ахроматическое и цветовое зрение. Анод и кальций, по этим данным, повышали чувствительность к зеленому, а катод и калий – к красному, т. е. симпатикотропные вещества действовали подобно аноду и Са, а парасимпатикотропные – подобно катоду и К. Эти факты имеют общность с фактами других авторов, полученными другими путями на других объектах. Так, Тинель (Tinel J. [135]) пишет: «Известно, что соли С а повышают тонус симпатической нервной системы... и увеличивают активность адреналина. Отсутствие кальция может устранить или даже извратить эффект адреналина... соли калия, наоборот, антагонистичны действию симпатического нерва: они повы- ----------------------------------------------------------- 92 ---------------------------------------------------------- шают тонус блуждающего нерва и эффект ацетилхолина... Можно сказать, что кальций действует как симпатический нерв, а калий, как блуждающий» '. Это же в итоге анализа ряда физиологических функций подчеркивают Цондак (Zondak S. [137]) и другие авторы. Кроме того, известно, что при отсутствии ионов не подействует и раздражение соответствующего нерва. «Ионы – орудия, посредством которых нерв осуществляет свою функцию» 2. В том же направлении действуют ионы натрия и магния: первый – по типу калия и катода, магний – по типу кальция и анода [68], [69], [70]. Таким образом, можно сказать, что порогу возбудимости зелено-и синеощущающего аппарата глаза соответствует меньшая величина в котором в числителе стоят (К) -f- (Na) отношения, -1 – : – - – -, а в знаменателе (Са) + (Mg) а порогу возбудимости красноощущающего аппарата – большая величина этого отношения. Опыты Поликаниной [|69J, [70] показали, что в гоматропинизирован-ном глазу почти незаметны реакции цветоощущающего аппарата на ионофорез Na- и Mg-ионов. Из фармакологии известно, что гоматропин препятствует соединению ацетилхолина с химическим веществом рецептивной субстанции эффекторной клетки. Эти опыты показывают, что в цветоощущающих клетках изменение содержания ацетилхолина перестает иметь значение, если в глаз введен гоматропин. Эти опыты подтверждают подобные наблюдения на ангиоскотомах и слепом пятне, впервые описанные Ро-славцевым [73], [74]. Поликаниной показано также, что звуковое центральное раздражение, повышавшее чувствительность к зеленому, сводится на нет идущим с периферии влиянием натрия (подобно только что описанным нашим опытам с нейтрализацией католического угнетения возбуждением второй сигнальной системы). Таким образом, вновь наблюдается отчетливое взаимодействие центральных и периферических раздражений. Если мы внимательно всмотримся в ~ -образную кривую реактивности (Кравкова), выражающую спектральную чувствительность к побочным раздражениям, то увидим в ней 3 неподвижные точки, это крайне фиолетовая 430-420 тр, крайне красная 680-700 тр и средняя желтая около 570 т\>. (рис. 3). Реактивность на раздражение имеет место только там, где затрагивается зеленоощущающий аппарат глаза, который занимает в нашем цветовом зрении особое положение. Приемником побочных раздражений является преимущественно он. В желтой точке нейтрализуется возбуждение зеленого и угнетение оранжево-красного – здесь ординаты для кривых возбуждения красного и зеленого равны, поэтому эта точка неподвижна. Особенность крайне красной точки – ее полная инертность в ответ на раздражение – зависит от отсутствия в крайних лучах спектра зеленого возбуждения, так же как и в области 430-420 тр. По мысли С. В. Кравкова, изменение возбужденного зеленоощу-щающего аппарата, связанное с повышением или понижением его возбудимости, оказывает влияние на возбудимость остальных двух цветоощущающих аппаратов нашего зрения: синего и красного. Это экспериментально подтверждено в опытах Медведевой (66). Она исследовала реакцию крайне красно- и фиолетовоощущаю-щего аппарата глаза при пропускании через глаз постоянного тока в 1 С. В. Кравков. Цветовое зрение. Изд-во АН СССР, 1951, стр. 141. 2 Там же. ----------------------------------------------------------- 93 ----------------------------------------------------------
Попытка подойти к разрешению вопроса о том, где же именно – в коре или в подкорке – протекает по преимуществу взаимодействие цве-тоощущающих аппаратов, была сделана Л. А. Шварц [108]. Она испытывала действие возбудителей преимущественно коркового – кофеина и преимущественно подкоркового – кордиамина и угнетателей преимущественно коркового – хлоралгидрата и преимущественно подкоркового – веронала на чувствительность к излучениям с длинами волн 435, 460, 520, 570, 625 и 700 тц. Автор показал, что кофеин повышал чувствительность ко всем длинам волн, а хлоралгидрат, наоборот, понижал чувствительность ко всем длинам волн. Подкорковые же фармакологические препараты на чувствительность к коротко- и длинноволновым излучениям оказывали противоположное действие, стимулятор кордиамин понижал чувствительность в области длинноволновых излучений и повышал в области коротковолновых, угнетатель – веронал оказывал противоположное действие. Отсюда С. В. Кравков делает вывод, что «физиологические механизмы, обусловливающие антагонистическую связь зеленоощущающего аппарата с красноощущающим, лежат, по-видимому, в подкорке» '. До сих пор мы рассматривали различные влияния на цветоощущаю-щий аппарат органа зрения. Но возбуждение цветоощущающего аппарата не только расширяет наши познания об окружающем мире, но и перестраивает физиологическое состояние организма и влияет на некоторые его реакции. В связи с нашими данными [82] о противоположном влиянии световой и темновой адаптации на кат- и анэлектротонический эффект в отношении электрической чувствительности глаза С. В. Кравковым и Р. Б. Зарецкой было проведено исследование ан- и катэлектротона на чувствительность к зеленому и красному цвету в темноте и на свету. В большинстве опытов (27 из 40) анэлектротон повышал чувствительность глаза к лучам с X =520 т\>- в темноте и понижал ее на свету. В отношении красных лучей чувствительность также изменялась противоположным образом на свету по сравнению с темнотой под влиянием поляризующего тока. Изменение «физиологического фона» при помощи цветовых раздражителей наблюдалось и в изменении внутриглазного давления, которое, по Р. Б. Зарецкой [35], снижалось в зеленом свете и повышалось в красном. Эти данные были использованы в глазной клинике при изучении и лечении глаукомы (в работах Р. Б. Зарецкой, Р. И. Бронштейн, А. В. Рославцева, Т. А. Алексидзе). Р. Б. Зарецкой [36] было показано, что слепое пятно сужается в зеленом свете и расширяется в красном. А. В. Рославцевым [74] наблюдалось сужение в зеленом свете и расширение в красном ангиоско^ом, т. е. тех мелких слепых мест в поле зрения, которые обусловливаются тенями кровеносных сосудов, находящихся перед светочувствительным слоем сетчатки. Зеленый свет – антагонист катоду – мог снимать действие последнего на слепое пятно, в зеленом свете от катода слепое пятно не расширялось (Зарецкая). Е. Н. Семеновской [92] показано повышение электрической чувствительности в зелено-желтых лучах в большей степени, чем в синих и красных. В крайне красных она даже часто снижалась, в то время как лабильность (критическая частота исчезновения фосфена) повышалась в красном и снижалась в зеленом свете. Данные эти подтверждены Р. Б. Зарецкой и в отношении электрической чувствительности – Г. К. Гуртовым. 1 С. В. Кравков. Цветовое зрение. Изд-во АН СССР, 1951, стр. 156. ----------------------------------------------------------- 95 ---------------------------------------------------------- По данным Л. А. Шварц [109], зеленый свет повышает слуховую чувствительность, а красный – понижает. В. К. Шеварева [110] нашла у всех испытуемых повышение мышечной работоспособности при зеленом освещении. С. В. Кравков, анализируя перечисленные выше результаты экспериментальных наблюдений, пришел к выводу, что, действуя на глаза, цветной свет действует на вегетативную нервную систему. Это следует из того, что физиологическая регуляция внутриглазного давления, границ слепого пятна, мышечной работоспособности и т. п. является функцией вегетативных, корковых и подкорковых центров, действующих на сосудистую систему. Действие же вегетативной нервной системы часто бывает подобно действию полюсов постоянного тока и определенным ионным сдвигам – преимущественно в соотношении ионов калия и кальция, необходимых для возбуждения симпатического и блуждающего нервов. Ионные же сдвиги связаны с изменением ацетилхолинового медиаторного обмена. В частности, зеленый свет в качестве симпатикотропного (как это следует из всего вышеизложенного) должен действовать в реагирующей системе в направлении относительного увеличения ионов кальция (и соответственно – уменьшения ацетилхолина). Красный же свет в качестве ваготропного должен увеличивать баланс калия (и соответственно ацетилхолина). Эти гипотетические положения С. В. Кравкова были подтверждены работами А. Я- Бунина [19], [20] и А. В. Рославцева [75]. А. Я. Бунин показал, что при адаптации к красному свету в сетчатках глаз лягушек ацетилхолина больше, чем при адаптации к зеленому свету. А. В. Рославцевым отмечено, что красный и зеленый свет не оказывает никакого действия на границы слепого пятна и ширину ангиоско-том, если глаза подверглись атропинизации. Это указывает на участие ацетилхолина в реакциях ангиоскотом и слепого пятна. А. В. Рославцев показал [75], что при потере крови (у доноров, у женщин в период mensis) реакции ангиоскотом извращаются: сужение их происходит не от зеленого, как в норме, а от красного света. Мы уже приводили выше мысль С. В. Кравкова, что объяснение описанным фактам следует искать в нарушении ионного равновесия (K) + (Na) aio отношение уменьшается под влиянием зеленого света и увеличивается под влиянием красного света. В 1956 г. в Неаполитанском журнале (Acta neurolog. Napoli, 11,3, 501 – 511) появился ряд интересных сообщений Парианте и Серра с соавторами, в которых авторы обнаружили снижение содержания калия в сыворотке крови и снижение активности холинестеразы при раздражении глаз человека белым и зеленым светом, а также уменьшение соотношения К/Са. При вспышках красного и желтого света наблюдалось обратное соотношение этих показателей. Авторы ссылаются на работу С. В. Кравкова и Л. П. Галочкиной [55], которая вероятно и явилась поводом для проведения подобного исследования. Таким образом, прямым биохимическим анализом крови была подтверждена гипотеза Кравкова о значении соотношения К/Са для объяснения противоположных реакций аппаратов, воспринимающих длинно- и коротковолновые излучения на побочные раздражители. В № 5 (стр. 865) те же авторы описывают наибольший гликемиче-ский эффект от красного и желтого света прямо пропорциональный длине волны, связывая его с торможением механизма регуляции содержания сахара в крови, который, как известно, связан с симпатической системой. Кравков же возбуждение красноощущающего аппарата связывал с возбуждением парасимпатической нервной системы, а зелено-ощущающего – с симпатической нервной системой. ----------------------------------------------------------- 96 ---------------------------------------------------------- В данном случае описанный процесс локализуется выше сетчатки, так как изменения изучавшихся функций происходили под влиянием освещения другого глаза. В доказательство этой мысли С. В. Кравков [48] ссылается на наши исследования [92], подтвержденные Г. К. Гуртовым (1950 [29]), об электрической чувствительности глаза ахроматов, т. е. полностью цвето-слепых, и ее изменений под влиянием раздражения их зрительного аппарата зеленым и красным светом. Как оказалось, электрическая чувствительность у ахроматов способна, так же как и у нормальных трихроматов, повышаться от зеленого света и понижаться от красного, даже если эти цветовые раздражители действовали на другой глаз. «Поскольку же коркового представительства цветного зрения в мозгу у ахроматов нет (и соответствующих цветовых ощущений они не имеют)... есть основание считать, что влияние цветности света сказывается здесь через посредство процессов, разыгрывающихся в подкорковой области головного мозга» (Кравков, 1952 [49]). Импульсы возбуждения при действии на те или иные цветоощу-щающие аппараты красного или зеленого цвета распространяются в различные области подкорки. Вполне вероятно, что у нормального трихромата «противоположные эффекты, вызываемые освещением глаз зеленым и красным светом, обусловливаются вегетативными реакциями противоположного знака, которые возникают в результате различного раздражения подкорки при действии на глаз света различного спектрального состава. Косвенным аргументом в пользу подобного понимания может явиться и то, что при некоторых заболеваниях, когда есть основание предполагать ненормальное состояние подкорки (как, например, при глаукоме), наблюдаются ненормальные реакции глаза на цветной свет» [49]. А. В. Рославцев показал, что ангиоскотомы у больных глаукомой в красном свете не расширяются, а сужаются, как при действии зеленого света, Е. Н. Семеновская и Л. А. Скворчевская наблюдали извращение реакции электрической чувствительности и лабильности у этих больных на красный свет [98]. Помимо теоретического значения, итоги работ С. В. Кравкова имеют и прямое практическое значение. Так, в глаукомном отделении Института им. Гельмгольца разрабатываются рациональные условия освещения, в том числе и зеленым светом, для больных глаукомой. Следует указать на еще одну важную проблему в области цветового зрения, которая разрабатывалась в лабораториях Кравкова. Это – проблема индукционных отношений внутри сетчатки и внутри зрительного анализатора между цветоощущающими аппаратами. В работе Л. П. Галочкиной [28] показано, что при реагирующей и индуцирующей точках зеленого, синего или фиолетового цвета положительная индукция (т. е. повышение чувствительности в реагирующем участке) почти отсутствует и, наоборот, сильно выражена отрицательная индукция (т. е. понижение чувствительности в реагирующем участке). Если же оба раздражителя оранжево-красного цвета, то сильнее выражена положительная индукция. В работе установлены многие важные и новые факты не только, для нормального трихроматического зрительного органа, но и для случаев с цветной аномалией. Отрицательная индукция выражена особенно сильно при бинокулярной индукции. Автор анализирует полученные результаты с теоретических позиций И. П. Павлова и Н. Е. Введенского. Л. И. Селецкая исследовала особенности красноощущающего аппарата при индукционных отношениях в зрительной системе [79]. Показана ----------------------------------------------------------- 97 ---------------------------------------------------------- ареактивность красноощущающего аппарата в области около 760 т\>. при индуцирующих цветах любой длины волн. Этот факт подтверждает особенность красноощущающего аппарата, описанную С. В. Кравковым. К работам, посвященным проблеме индукционных отношений, следует отнести и работу С. В. Кравкова и Е. Н. Семеновской [57], где показано, что после освещения макулярной области сетчатки красным или зеленым светом изменение возбудимости в периферическом палочковом аппарате обнаруживает сложные индукционные отношения между палочками, расположенными на 10 и на 40е к периферии. В то время как световая чувствительность после красного «засвета» повышается в области, лежащей на 10° к периферии, в области, лежащей на 40° к периферии, она понижается. После зеленого «засвета» наблюдаются обратные соотношения. Здесь подмечена и разная скорость распространения индукционного влияния – большая для красного и меньшая для зеленого света. Позднее Е. Н. Семеновской (1950 – 1954) показана разная лабильность в зрительной системе при освещении глаз красным и зеленым светом и сделано допущение, что в явлении индукционных отношений большую роль играет разная лабильность взаимновлияющих друг на друга систем.
Сенсорные условные рефлексы Большое место в трудах лаборатории С. В. Кравкова занимают исследования сенсорных условных рефлексов в зрительном анализаторе, начатые в 1936 г. Вот что писал С. В. Кравков об этих работах в статье «Учение И. П. Павлова об условных рефлексах в офтальмологии» («Вестник офтальмологии», № 6, 1949). «Учение Павлова об условных рефлексах, зародившееся во время специальных опытов с пищеварением, выросло в широкую физиологическую концепцию, являющуюся гордостью русской и мировой науки. Первоначально исследования условных рефлексов охватывали лишь реакции секреторного и двигательного характера. Однако Павлов писал: «Временная нервная связь есть универсальнейшее физиологическое явление в животном мире и в нас самих». Нет поэтому никакого основания полагать, что законы, открытые Павловым применительно к деятельности слюнной железы, не могут быть применены и при изучении зрительного анализатора». В 1936 г. были впервые описаны сенсорные условные рефлексы, конечным выражением которых являлась уже не секреторная или двигательная реакция, а определенные изменения возбудимости органов чувств (в лаборатории С. В. Кравкова А. И. Богословский, независимо от них А. И. Долин, а также Г. X. Кекчеев [33]). В работах А. И. Богословского (1936) было отмечено, что после ряда опытов со световым раздражением («засветом») одного глаза на определенной минуте темновой адаптации, которое всегда влечет за собой повышение чувствительности другого глаза к электрическому току, электрическая чувствительность глаза повышается на тех же минутах темновой адаптации и в последующих опытах, в которых никакого светового раздражения другого глаза (безусловного раздражителя) уже не применяется. Возникал условный рефлекс «на время», подобный тому, который в лаборатории Павлова описан Феокритовой (1912) применительно к деятельности слюнной железы. Наблюдавшиеся факты заставили автора провести специальные опыты по выработке условнорефлекторного изменения электрической чувствительности глаза. ----------------------------------------------------------- 98 ---------------------------------------------------------- Возникновение условных рефлексов на время отчетливо наблюдалось А. И. Богословским также на функции слияния световых мельканий при центральном и периферическом зрении. Условно-рефлекторное изменение остроты зрения получено М. А. Севрюгиной [76], А. И. Богословским [|5], [6], [12]; различительной чувствительности, световой чувствительности – В. И. Рождественской [72]. Все эти опыты показали, что сенсорные условные рефлексы глаза подчиняются закономерностям павловских условных рефлексов. При неподкреплении их безусловными раздражителями они угасают: можно вырабатывать дифференцировку одного условного раздражителя от другого. Было установлено, что сенсорные условные рефлексы у человека возникают быстрее, чем слюнные условные рефлексы у собаки. Достаточно бывает 1 – 7 сочетаний, чтобы образовался условный рефлекс. Вырабатываемые дифференцировки могут быть очень тонки (Богословский, 1940). Достаточно сказать, что электрическая чувствительность глаза может изменяться в ответ на условный раздражитель – стук метронома с частотой 120 раз в минуту и не изменяться при частоте 118 раз в минуту. Автор полагает, что в основе элементарного узнавания и различения по памяти лежит дифференцировочное торможение. После того как было обнаружено, что чувствительность глаза может изменяться по законам условных рефлексов, круг раздражителей, способных так или иначе влиять на зрение, можно считать в сущности безграничным. «Не нужно большого воображения, – писал Павлов, – чтобы сразу увидеть, какое прямо неисчислимое множество условных рефлексов постоянно практикуется сложнейшей системой человека, поставленной в часто широчайшей не только общеприродной среде, но и в специально-социальной среде» '. Если в силу законов временной связи любой раздражитель может оказывать влияние на зрение, естественно было ожидать, что подобный же эффект может наблюдаться и в том случае, если у нас возникает лишь достаточно яркий воображаемый образ (представление) действующего раздражителя. Ведь между таким представлением и наличием самого раздражителя всегда имелось совпадение во времени, т. е. в. нашей нервной системе существовала лежавшая в основе условного рефлекса временная связь. Выполненные впервые в лаборатории Кравкова работы (А. И. Богословский [6], [7], О. А. Добрякова [30], А. А. Дубинская, Е. Н. Семеновская [83]) показали, что, действительно, функции зрения (электрическая чувствительность глаза) могут изменяться и под влиянием лишь представлений, возникающих у испытуемого. Так, например, воображение того, что перед глазами находится большой освещенный экран, влекло за собой заметное повышение уровня электрической чувствительности глаза. После того как испытуемый переставал воображать светлый экран, чувствительность возвращалась к исходному уровню. Отмечено также, что не только представление тех или иных раздражителей может оказывать такое действие, но и произнесенное вслух название этих раздражителей. Такой эффект понижения электрической чувствительности наблюдался в опытах Добряковой [30], когда испытуемому показывали лишь написанные слова: «кислый, как лимон». Таким образом, мы видим, что чувствительность органов чувств и, в частности, зрения изменяется и при возбуждении второй сигнальной системы. В опытах Л. А. Шварц было выяснено, что у человека при использовании слов как условных раздражителей в нормальных условиях иг- 1 И. П. Павлов. Поли. собр. сочинений. Изд-во АН СССР, 1951, III, 2, 325..95 ---------------------------------------------------------- 100 ----------------------------------------------------------
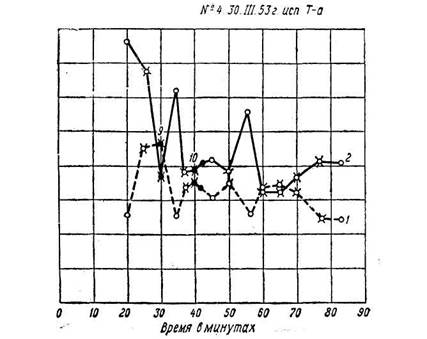
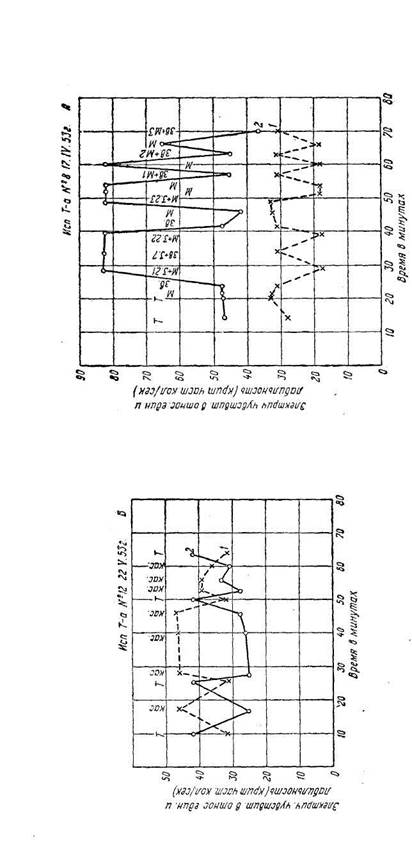
Эффект условного раздражителя мог угасать в течение одного опыта без подкрепления соответствующим светом, мог иногда снова возникать на другой день (рис. 5). Угасание условного значения метронома можно было ускорить, выработав, например, торможение на звонок. Если же метроном сочетался с цветовым раздражителем длительно (10 – 20 мин.), то его условное действие могло длиться месяцами. Наблюдалась хорошо выраженная дифференцировка метрономов, сочетавшихся с разными цветами. Условный раздражитель может воспроизводить процесс, протекавший во времени, а не одномоментное состояние зрительного анализатора. Описанной методикой можно было определять типологические особенности высшей нервной деятельности, но в данной работе мы не могли этим заниматься. Переделка метрономов оказалась, как этого и нужно было ждать, гораздо более трудной работой. При переделке М-120, сочетавшегося с красным светом и вызывавшего «красную» реакцию электрической чувствительности и лабильности, на «зеленый лад» путем сочетания его с зеленым светом потребовалось вместо 1-3 сочетаний 12-15 сочетаний, причем довольно длительно тянулся период нулевых реакций, когда метроном не вызывал никаких сдвигов ни электрической чувствительности, ни лабильности. Но когда переделка удалась и М-120 стал вызывать «зеленую» реакцию обоих параметров, то обнаружилось, что у испытуемого сам безусловный красный свет стал действовать как зеленый, т. е. стал понижать лабильность и повышать электрическую возбудимость оптического анализатора. Потребовалось много месяцев, чтобы вернуть реакцию на красный свет к нормальному ее виду. У этой испытуемой, как по нашим данным, так и по данным других экспериментаторов, работавших по другим методикам, возбудительный процесс отличался большой стойкостью [100]. Краткие сочетания условного раздражителя с безусловным приводили к угашению рефлекса в тот же день в течение опыта, после того как он продержался 30 – 40 мин. Однако иногда на другой день в начале опыта рефлекс мог возникнуть снова. Описанные опыты устанавливают, что в механизме физиологической противоположности красно- и зеленоощущающих аппаратов, открытой С. В. Кравковым, и в механизме индукционных отношений принимают обязательное участие противоположные сдвиги электрической чувствительности и лабильности зрительного анализатора, которые выражают собой фазность процессов, возникающих в зрительной системе при адаптации глаз к красному и зеленому свету, и связанные, по-видимому, с индукционными отношениями между красно- и зеленоощущающими аппаратами. Краткость его делает очень удобным применение этого метода во всех тех случаях, где длительность опыта или утомительна для испытуемого, как, например, в клинике, или может нивелировать действие изучаемой обстановки на человека, что может произойти при изучении физиологического состояния человека в условиях производства, спорта и т. п. На рисунках 4 и 5 показаны некоторые протоколы наших опытов с условными рефлексами. В них, как нам кажется, особый интерес представляет тот факт, что условный раздражитель в сенсорной области вызывает целую серию сдвигов различных функций. Эти сдвиги могут быть взаимно противоположно направлены, как, например, электрическая чувствительность и лабильность, точно так же, как это имеет место при действии безусловного раздражителя. А. В. Рославцевым [73], [74] показаны очень интересные наблюдения об условнорефлекторных изменениях ангиоскотом и слепого пятна, а также внутриглазного давления (последнее наблюдала и Р. И. Бронштейн [18]). ---------------------------------------------------------- 101 ----------------------------------------------------------
Автор ранее наблюдал их расширение при действии красного света. Оказывается, что если сочетать красный свет со звуком мотора, то после очень небольшого количества сочетаний происходит расширение ангиоскотом и слепого пятна на один звук мотора. На звук другого мотора другой тональности возникает дифференцировка и рефлекс не возникает. Он, как и все условные рефлексы, затухает при отсутствии подкрепления. Нам остается сказать несколько слов еще об одном большом разделе работ, проводившихся в лаборатории С. В. Кравкова, – об электрофизиологических исследованиях зрительной системы в более широком смысле, чем электрическая чувствительность и действие поляризующего тока.
|
||
|
|
Последнее изменение этой страницы: 2021-04-20; просмотров: 159; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.119 (0.018 с.) |