
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Бета-Окисление жирных кислот. Последовательность реакций. Энергетическое значение.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
1. Жирные кислоты, как и глюкоза, являются основными «топливными молекулами». Большинство тканей, кроме нервной ткани, эритроцитов (в которых отсутствуют митохондрии), использует жирные кислоты как источник энергии. Жирные кислоты, проникающие из крови в клетку, сначала подвергаются реакции активации под действием фермента ацил-КоА-синтетазы:
2. β-Окисление жирных кислот - это специфический путь катаболизма жирных кислот, продуктом которого является ацетил-КоА. Название «β-окисление» эти реакции получили потому, что окисление в радикале жирной кислоты происходит по β-углеродному атому. β-Окисление жирных кислот и последующее за ним окисление ацетил-КоА в ЦТК служат источником энергии для синтеза АТФ. Процесс β-окисления происходит в матриксе митохондрий и только в аэробных условиях, так как он связан с ЦПЭ через коферменты дегидрогеназ, водород от которых поступает в ЦПЭ. Внутренняя мембрана митохондрий непроницаема для ацил-КоА, поэтому существует система переноса радикала жирных кислот через мембрану в комплексе с молекулой карнитина (рис. 8.18). Фермент карнитинацилтрансфераза I, осуществляющий перенос ацила на карнитин на внешней мембране митохондрий, является регуляторным в процессе β-окисления, так как определяет скорость переноса жирных кислот внутрь митохондрий.
Во внешней мембране митохондрий находится фермент карнитинацилтрансфераза I, который катализирует перенос ацила с КоА на небольшую молекулу - карнитин. Затем ацилкарнитин с помощью транслоказы проходит через внутреннюю мембрану митохондрий, где фермент карнитинацилтрансфераза II переносит ацил на внутримитохондриальный HS-KoA. Фермент карнитинацилтрансфераза I - регуляторный в процессе β-окисления, ингибитором этого фермента является малонил-КоА 3. После того как ацил-КоА попадает в матрикс митохондрий, начинается процесс β-окисления, представляющий собой четыре последовательные реакции, которые заканчиваются укорочением жирной кислоты на два углеродных атома, отделяющиеся в форме ацетил-КоА (рис. 8.19). Эти четыре реакции β-окисления (дегидрирование, гидратация, дегидрирование, отщепление ацетил-КоА) обычно называют «циклом β-окисления», так как имеется в виду, что одни и те же реакции повторяются с радикалом жирной кислоты до тех пор, пока вся кислота не превратится в ацетильные остатки. Количество молекул АТФ, которые образуются при окислении жирной кислоты, можно рассчитать по формуле:
• состояния голодания или сытости (т.е. соотношения инсулин - глюкагон); • активности регуляторного фермента карнитинацилтрансферазы I; • доступности субстрата - жирных кислот; • потребности клетки в энергии; • доступности кислорода. Под действием инсулина, гормона «сытого состояния» в клетках печени появляется ингибитор регуляторного фермента карнитинацилтрансферазы I - малонил-КоА (рис. 8.18). Это вещество образуется в первой реакции синтеза жирных кислот, катализируемой регуляторным ферментом синтеза жирных кислот - ацетил-КоА-карбоксилазой. Появление в гепатоцитах малонил-КоА немедленно ингибирует β-окисление жирных кислот, таким образом, синтез и окисление жирных кислот не могут происходить одновременно. При голодании или физической работе под действием гормонов глюкагона или адреналина синтез малонил-КоА снижается и скорость β-окисления увеличивается, следовательно, окисление жирных кислот становится важным источником энергии при голодании или длительной физической работе. В мышцах карнитинацилтрансфераза I также ингибируется малонилКоА. Хотя эта ткань не синтезирует жирные кислоты, в ней имеется изофермент ацетил-КоА-карбоксилазы, синтезирующий малонил-КоА для регуляции β-окисления. Этот изофермент фосфорилируется под действием протеинкиназы А, активируемой в клетках адреналином, и АМФ-зависимой протеинкиназой и таким образом ингибируется; концентрация малонилКоА снижается. Следовательно, при физической работе, когда в клетке появляется АМФ, под действием адреналина активируется β-окисление, однако его скорость зависит еще и от доступности кислорода. Поэтому β-окисление становится источником энергии для мышц только через 10-20 минут после начала физической нагрузки (так называемые аэробные нагрузки), когда приток кислорода к тканям увеличивается. 5. При голодании и физической работе активируется липолиз в жировой ткани и поток жирных кислот в ткани увеличивается. Жирные кислоты становятся важным источником энергии для таких тканей, как скелетные мышцы, миокард, печень. Однако мозг не может использовать жирные кислоты как источник энергии, потому что они не проникают через гематоэнцефалический барьер, являясь гидрофобными молекулами. Поэтому в таких ситуациях, особенно при длительном голодании, печень перерабатывает ~50% поступающих в нее жирных кислот в другие источники энергии - кетоновые тела, которые может утилизировать нервная ткань. 56.Связь окисления жирных кислот с цитратным циклом и дыхательной цепью. Физиологическое значение.
Оба этапа сопряжены с дыхательной цепью. b -окисление происходит в митохондриях клетки, а жирная кислота поступает из кровотока в цитозоль, где активируется путем конденсации с коферментом А, образуя тиоэфир.
Для того чтобы пройти через митохондриальную мембрану, жирной кислоте необходим специальный переносчик - карнитин. Поэтому сначала ацильная группа переносится с ацил-СоА на карнитин. Ацил-карнитин пересекает мембрану и отдает свой ацильный фрагмент снова на кофермент А. Ферментом этой обратимой реакции служит ацил-карнитинтрансфераза. Результатом четырех последующих реакций b -окисления является отщепление двухуглеродного фрагмента и перенос его на кофермент А с образованием ацетил-СоА, который затем может включаться в цитратный цикл для полного окисления. Укороченная ацильная цепь вторично входит в цикл b -окисления. Конечным итогом повторяющихся циклов b -окисления будет окисление всей ацильной цепи до ацетил-СоА.
|
||||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 633; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.155.253 (0.007 с.) |

 Жирные кислоты могут вступать в различные реакции: окисления, синтеза ТАГ или фосфолипидов только в виде КоА-производных.
Жирные кислоты могут вступать в различные реакции: окисления, синтеза ТАГ или фосфолипидов только в виде КоА-производных.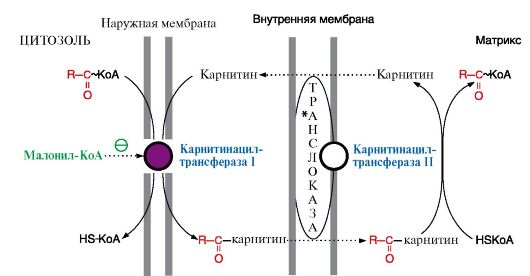 Рис. 8.18. Транспорт высших жирных кислот через мембраны митохондрий
Рис. 8.18. Транспорт высших жирных кислот через мембраны митохондрий 2* - энергия двух макроэргических связей АТФ используется на активацию жирной кислоты.
2* - энергия двух макроэргических связей АТФ используется на активацию жирной кислоты.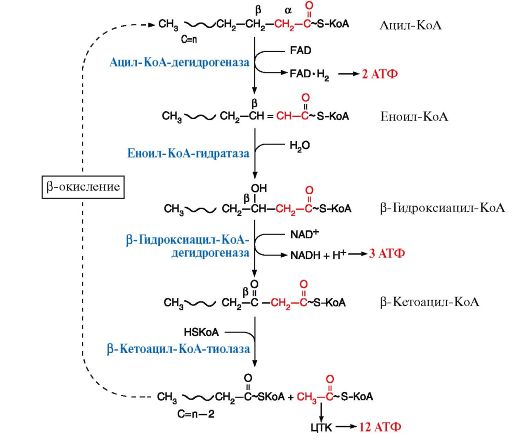 Рис. 8.19. Реакции β-окисления жирных кислот
Рис. 8.19. Реакции β-окисления жирных кислот 4. Регуляция β-окисления. Скорость процесса β-окисления зависит от ряда факторов:
4. Регуляция β-окисления. Скорость процесса β-окисления зависит от ряда факторов: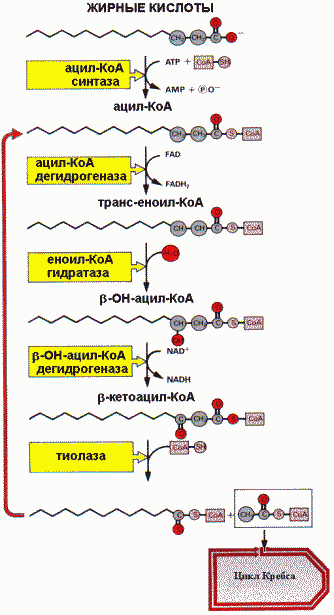 Окисление жирных кислот
Окисление жирных кислот



