
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Регуляция ферментов путем их фосфорилирования — дефосфорилирования
Протеинкиназы катализируют фосфорилирование белков по гидроксильным группам серина, треонина и тирозина. Если фосфорилируемые белки это тоже ферменты, то их активность в результате фосфорилирования в одних случаях уменьшается, в других — увеличивается. Например, в клетках жировой ткани есть липаза, существующая в двух формах — фосфопротеина и простого белка.
Регуляция ферментов. Эти формы могут превращаться друг в друга. Фосфопротеин образуется в результате действия протеинкиназы и может вновь превращаться в простой белок при действии фосфопротеинфосфатазы — фермента, гидролитически отщепляющего фосфорную кислоту от фосфопротеинов Фосфорилированная липаза обладает значительно более высокой активностью, чем нефосфорилированная. Первичная, вторичная и третичная структуры ДНК. Химические связи. Репликация ДНК. Первичная структура нуклеиновых кислот (НК) - это порядок чередования нуклеотидов в полинуклеотидной цепи, связанных между собой 3',5'-фосфодиэфирной связью. Образующиеся полимеры имеют фосфатный остаток на 5'-конце и свободную -ОН-группу пентозы на 3'-конце (рис. 3.1). Штрихами обозначают углеродные атомы пентозы для того, чтобы отличать их от атомов, входящих в азотистые основания.
Для краткого изображения последовательности нуклеотидов в нуклеиновых кислотах пользуются однобуквенным кодом. При этом запись осуществляют слева направо таким образом, что первый нуклеотид имеет свободный 5'-фосфатный конец, а последний -ОН-группу в 3'-положении рибозы или дезоксирибозы.
Рис. 3.1. Первичная структура нуклеиновых кислот. Х = Н для ДНК, Х = ОН для РНК Связи в молекуле нуклеиновых кислот: 1 - 5'-фосфоэфирная; 2 - N-гликозидная; 3 - 3',5'-фосфодиэфирная 2. Пространственная структура ДНК. Вторичная структура представляет собой правозакрученную спираль (рис. 3.2), в которой две полинуклеотидные цепи расположены антипараллельно и удерживаются относительно друг друга за счет водородных связей между комплементарными азотистыми основаниями: А = Т и G = С. Цепи молекулы ДНК не идентичны, но комплементарны друг другу: если известна первичная структура одной цепи, то последовательность нуклеотидов другой цепи задается правилом комплементарности оснований: Т одной цепи соответствует А, а С - G в другой цепи. Поэтому в молекуле ДНК количество адениловых нуклеотидов равно количеству тимидиловых нуклеотидов (А = Т), а количество гуаниловых равно количеству цитидиловых нуклеотидов (G = С). Соотношение А + Т / G + С - величина постоянная и является видоспецифической характеристикой организма. Основания нуклеотидов обращены внутрь молекулы и лежат в одной плоскости, которая практически перпендикулярна оси спирали. Между основаниями, расположенными друг под другом, возникают гидрофобные взаимодействия. Дезоксирибозофосфатные остатки образуют остов спирали. На один виток спирали приходится 10 нуклеотидных пар. Третичная структура ДНК формируется в результате ее взаимодействия с белками. Каждая молекула ДНК упакована в отдельную хромосому, в составе которой разнообразные белки связываются с отдельными участками ДНК и обеспечивают суперспирализацию и компактизацию молекулы. В период покоя комплексы ДНК с белками распределены равномерно по объему ядра, образуя хроматин. Белки хроматина включают две группы: гистоны и негистоновые белки.
Гистоны - небольшие белки с молекулярной массой от 11 000 до 22 000 Д и высоким содержанием лизина и аргинина. Четыре типа гистонов в количестве восьми молекул (по две каждого вида) образуют комплекс - нуклеосомный кор. Этот комплекс за счет ионных связей взаимодействует с отрицательно заряженными фосфатными группами участка ДНК длиной около 146 нуклеотидных пар (примерно 1,75 витка вокруг кора) и образует структуру, называемую нуклеосомой. Между нуклеосомами находятся участки ДНК длиной около 30 нуклеотидных пар - линкерные участки, к которым присоединяются молекулы гистона Н1 (рис. 3.3). Негистоновые белки представлены множеством ферментов и белков, участвующих в синтезе ДНК, РНК, регуляции этих процессов и компактизации ДНК.
Репликация - матричный процесс. Во время репликации каждая из двух цепей ДНК служит матрицей для образования новой цепи. Субстратами и источниками энергии для синтеза ДНК являются дезоксирибонуклеозидтрифосфаты (дНТФ: дАТФ, дГТФ, дЦТФ, дТТФ).
|
|||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 127; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.234.62 (0.004 с.) |

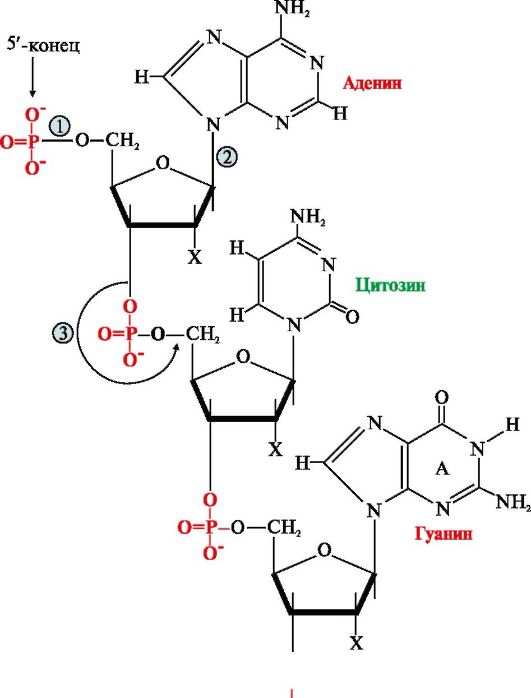
 Рис. 3.2. Двойная спираль ДНК.
Рис. 3.2. Двойная спираль ДНК.


