

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Процесс включает следующие основные этапы.
Содержание книги
- Биологические функции белков. Взаимодействие с лигандами.
- Конформация белковых молекул (вторичная и третичная структуры). Активный центр белков. Связывание лигандов.
- Основы кинетики ферментативного катализа
- Кофакторы ферментов. Коферментные функции витаминов группы В.
- Ингибиторы активности ферментов
- Классификация и номенклатура ферментов
- Наследственные (первичные) энзимопатии: нарушение обмена при алкаптонурии, фенилкетонурии, гипераммониемии.
- Активаторы и ингибиторы ферментов
- Регуляция ферментов путем их фосфорилирования — дефосфорилирования
- Процесс включает следующие основные этапы.
- III. Исключение праймеров. Завершение формирования отстающей цепи ДНК
- Биосинтез рнк (транскрипция). Посттранскрипционные модификации рнк
- Биосинтез белков. Основные компоненты и этапы этого процесса. Посттрансляционный процессинг белков.
- Регуляция экспрессии генов. Теория оперона. Индукция и репрессия синтеза белков.
- Молекулярные механизмы генетической изменчивости. Мутации. Наследственные протеинопатии.
- Молекулярные механизмы генных, хромосомных и геномных мутаций
- Биологический код, его свойства, значение в биосинтезе белка. Взаимодействие кодонов с антикодонами
- Макроэргические соединения. Их роль в клетке
- Характеристика высокоэнергетических фосфатов. Цикл атф-адф
- Тема 5. 6. Разобщение дыхания и синтеза атф
- Глк-глюкоза, Фру-фруктоза, Гал-галактоза
- Синтез глюкозы (глюконеогенез)
- Биосинтез и мобилизация гликогена: последовательность реакций, регуляция гормонами, физиологическое значение.
- Регуляция активности фосфорилазы гормонами.
- Основные пути превращения глюкозы в печени.
- Регуляция обмена углеводов инсулином, глюкагоном, адреналином, кортизолом.
- Пентозофосфатный путь окисления глюкозы, физиологическое значение.
- Гликолипиды и гликопротеины. Представления о строении и функциях их углеводных компонентов.
- I. Структура, классификация и свойства основных липидов организма человека
- Б. Структура и классификация фосфолипидов и сфинголипидов
- Пищевые жиры: норма суточного потребления, переваривание, всасывание продуктов.
- В поддержании гомеостаза холестерола в организме. Биохимия желчнокаменной болезни
- Тема 8. 3. Хиломикроны - транспортная форма экзогенных жиров
- Бета-Окисление жирных кислот. Последовательность реакций. Энергетическое значение.
- Механизмы биосинтеза жирных кислот. Регуляция этого процесса.
- Регуляция синтеза жирных кислот.
- Биосинтез жиров в печени и жировой ткани. Регуляция синтеза жиров
- Депонирование и мобилизация жиров в жировой ткани. Механизм регуляции активности липазы гормонами.
- Фосфолипиды, строение, биологическая роль.
- Холестерин, этапы биосинтеза, биологические функции, регуляция биосинтеза.
- Транспортные формы холестерина. Нарушения транспорта. Семейная гиперхолестеринемия. Атеросклероз.
- Причины и факторы риска развития атеросклероза
- Первичные эндогенные формы истощения
- Представления о биосинтезе фосфолипидов. Липотропные вещества.
- Связь между обменом белков и углеводов
- Влияние инсулина, глюкагона и адреналина на обмен жиров.
- Тема 9. 2. Переваривание белков в желудке и кишечнике, всасывание аминокислот
- Реакции трансаминирования, ферменты, их коферментная группа. Биологическое значение реакций. Определение аминотрансфераз с диагностической целью
- Окислительное дезаминирование аминокислот. Глутаматдегидрогеназа. Значение этой реакции.
- Непрямое дезаминирование аминокислот: последовательность реакций, ферменты, биологическое значение.
I. Формирование репликативной вилки.
II. Синтез новых цепей ДНК.
III. Исключение праймеров. Завершение формирования отстающей цепи
ДНК.
I. Формирование репликативной вилки (рис. 3.5) идет при участии:
ДНК-топоизомеразы, которая является «обратимой нуклеазой». Сначала фермент разрывает 3'-,5'-фосфодиэфирную связь в одной из цепей ДНК
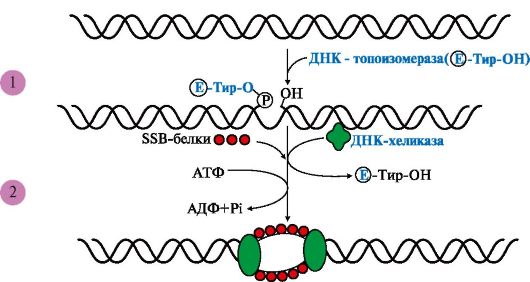 Рис. 3.5 Участие ДНК-топоизомеразы в образовании репликативной вилки. Рис. 3.5 Участие ДНК-топоизомеразы в образовании репликативной вилки.
1 - фермент расщепляет 3',5'-фосфодиэфирную связь в одной из цепей ДНК и присоединяется к 5'-концу в точке разрыва; 2 - в область разрыва цепи присоединяются две молекулы ДНК-хеликаз и обеспечивают локальное разделение двойной спирали ДНК. ДНК-топоизомераза восстанавливает расщепленную 3',5'-фосфодиэфирную связь и отделяется, а к одноцепочечным участкам присоединяются SSB белки
и присоединяется к 5'-концу в точке разрыва, вызывая сброс суперспиралей ДНК-хроматина. Это облегчает присоединение в область разрыва цепи двух молекул ДНК-хеликаз и образование репликативной вилки. По окончании формирования репликативной вилки ДНК-топоизомераза восстанавливает целостность молекулы ДНК и отделяется;
ДНК-хеликаз - ДНК-зависимых АТФаз, использующих энергию АТФ для расплетения двойной спирали ДНК;
SSB (single strand binding)- белков, связывающихся с одноцепочечными участками ДНК. Эти белки, не закрывая оснований, предотвращают повторное комплементарное скручивание матричных цепей и образование шпилек.
II. Синтез новых цепей ДНК
На этой стадии дочерние нити ДНК образуются на обеих нитях материнской ДНК. Процесс катализирует несколько ДНК-полимераз, которые синтезируют полинуклеотидные цепи из дНТФ: дАТФ, дГТФ, дТТФ и дЦТФ в направлении от 5'- к 3'-концу антипараллельно матрице, имеющей направление от 3'- к 5'-концу (рис. 3.6).
Новые цепи синтезируются по-разному. На матрице ДНК с направлением от 3'- к 5'-концу цепь растет непрерывно по ходу движения репликативной вилки и называется лидирующей. На матрице с направлением от 3'- к 5'-концу вторая цепь синтезируется против движения репликативной вилки в виде коротких отрезков - фрагментов Оказаки. Рост этой цепи начинается только тогда, когда на матрице ДНК появляется одноцепочечный участок длиной около 200 нуклеотидов, поэтому ее называют запаздывающей или отстающей.
 Рис. 3.6. Рост новых цепей в области репликативной вилки. Рис. 3.6. Рост новых цепей в области репликативной вилки.
Лидирующая нить растет непрерывно, а отстающая - в виде фрагментов Оказаки, каждый из которых включает:
- РНК-праймер (~10 нуклеотидов);
- участок ДНК, примерно равный длине цепи из 150 нуклеотидов
ДНК-полимеразы δ, β и ε не способны инициировать синтез новых цепей ДНК, они могут лишь удлинять имеющуюся нуклеотидную цепь. Синтез лидирующей и отстающей нитей начинается с образования затравки или праймера -олигорибонуклеотида (РНК), включающего около 10 мононуклеотидов. Его образование катализирует праймаза - субъединица ДНК-полимеразы α. Далее этот же фермент, используя в качестве субстратов дНТФ, переключается на образование ДНК и включает во вновь синтезируемую нить 20-50 дезоксирибонуклеотидов, после чего заменяется другими ДНК-полимеразами. Синтез лидирующей цепи продолжает ДНК-полимераза δ, а отстающей - ДНК-полимераза δ или ε. Оба фермента, помимо, полимеразной обладают еще и экзонуклеазной активностью. В ходе синтеза они могут исправлять допущенную ошибку и отщеплять неправильно включенный нуклеотид, что обеспечивает высокую точность синтеза ДНК.
|




