
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Регуляция экспрессии генов. Теория оперона. Индукция и репрессия синтеза белков.Содержание книги
Поиск на нашем сайте
Адаптация организмов к различным воздействиям окружающей среды часто осуществляется путем изменения экспрессии (активности) генов, т.е. изменения скорости транскрипции определенных участков на молекулах ДНК. Этот процесс, в деталях изученный на бактериях, включает взаимодействие специфических белков с участками ДНК в непосредственной близости от стартового участка транскрипции - промотора. 1. Адаптивная регуляция активности генов у прокариотов получила объяснение в теории оперона. Согласно этой теории на молекуле ДНК прокариотов присутствуют определенные участки - опероны. В состав этих участков ДНК входят структурные гены, содержащие информацию о группе функционально взаимосвязанных белков, которые участвуют в одном и том же метаболическом пути, промотор и оператор. Участки промотора и оператора частично перекрываются. Транскрипцию структурных генов контролирует оператор, присоединение к которому белка-репрессора не позволяет РНК-полимеразе связаться с промотором и начать транскрипцию. Белок-репрессор синтезируется в клетке с постоянной скоростью, его строение кодирует мРНК, транскрибируемая с гена-регулятора, расположенного на некотором расстоянии от оперона, работу которого контролирует его белковый продукт. Если оперон регулируется по механизму индукции (например, лактозный оперон), то в отсутствии индуктора (лактозы) белок-репрессор связан с оператором. Комплекс белок-репрессор-оператор препятствует связыванию РНК-полимеразы с промотором, и транскрипция структурных генов оперона не идет. Когда концентрация индуктора в клетке возрастет, то он присоединится к белку-репрессору, который имеет центр для связывания индуктора, изменяет его конформацию и снижает сродство к оператору. Комплекс индуктор-белок-репрессор теряет сродство к ДНК и уходит в цитозоль клетки. РНК-полимераза связывается с промотором и транскрибирует структурные гены. Идет синтез белков, закодированных в данном опероне (рис. 3.19). При регуляции оперона по механизму репрессии (например, гистидиновый или триптофановый опероны) белок-репрессор, постоянно синтезируемый в клетках, не имеет сродства к оператору. Когда к белку-репрессору присоединится небольшая молекула - корепрессор (гистидин или триптофан, как правило, конечный продукт метаболического пути, ферменты которого закодированы в структурных генах оперона), то в результате конформационных изменений комплекс белка-репрессора с корепрессором приобретает сродство к оператору и прекращает транскрипцию. Так, гистидиновый оперон содержит 10 структурных генов, кодирующих строение ферментов синтеза гистидина. Добавки гистидина в среду выращивания клеток E. coli вызывают снижение количества, а затем и полное исчезновение из внутриклеточного содержимого ферментов синтеза гистидина (рис. 3.20). 2. Адаптивная регуляция активности генов у эукариотов обеспечивает изменения скорости транскрипции отдельных генов в ответ на меняющиеся условия внутренней и внешней среды. В клетках многоклеточных организмов часть генов кодирует белки «домашнего хозяйства», которые синтезируются с постоянной скоростью и обеспечивают жизнеспособность клеток. Это - гены ферментов, участвующие в биологическом окислении, синтезе АТФ, образовании компонентов мембран и т.д. Регуляция у высших организмов отличается от регуляции транскрипции у прокариотов многообразием сигналов, которые контролируют не только начало процесса на молекуле ДНК, но и частоту, с которой он
А - в отсутствие индуктора в среде белок-репрессор связывается с оператором. РНК-полимераза не может присоединиться к промотору, транскрипция не идет; Б - в присутствии индуктора белок-репрессор образует комплекс с молекулами индуктора, меняет конформацию и теряет сродство к оператору. РНК-полимераза транскрибирует гены А, В, С и происходит синтез белков: β-галактозидазы, пермиазы, галактозидтрансацетилазы, участвующих в утилизации лактозы
А - в отсутствие корепрессора белок-репрессор неактивен и не имеет сродства к оператору до тех пор, пока небольшая молекула-корепрессор не свяжется с ним; Б - в присутствии корепрессора комплекс белок-репрессор-корепрессор связывается с оператором и прекращает транскрипцию происходит (рис. 3.21). ТАТА-участок промотора присоединяет ТАТА-связывающий белок, факторы транскрипции обеспечивают взаимодействие с РНК-полимеразой и определяют стартовую точку транскрипции. Минимальный синтез мРНК становится возможным после связывания РНК-полимеразы с группой дополнительных транскрипционных факторов. Если кроме указанных компонентов с РНК-полимеразным комплексом связываются белки, присоединенные к регуляторным участкам ДНК, то скорость транскрипции меняется. Она возрастает, если это будут белки, взаимодействующие с участками ДНК -энхансерами (усилителями), и снижается, если к транскрипционному комплексу
Область, обеспечивающая регуляцию экспрессии генов, включает в себя промоторный участок и дополнительные регуляторные последовательности, в которые входят энхансеры, сайленсеры, гормон-чувствительные участки, последовательности GC, СААТ и др. Белки, связывающиеся с ДНК в этих участках, называют специфическими регуляторными белками. Они влияют на скорость транскрипции генов, взаимодействуя с белками-посредниками или коактиваторами, передающими сигнал на основные транскрипционные факторы и РНК-полимеразу: 1 - регуляторные участки ДНК; 2 - регуляторные белки; 3 - белки-коактиваторы; 4 - РНК-полимеразный комплекс присоединится белок, связывающийся с участком сайленсера (тушитель транскрипции). Регуляторные зоны ДНК - энхансеры и сайленсеры, гормон-чувствительные участки, специфические регуляторные последовательности (например, GC, CAAT и др.) различны по числу и расположению на молекуле ДНК для разных генов в разных тканях, т.е. являются тканеспецифическими характеристиками. Они могут располагаться за тысячи нуклеотидных пар от стартовой точки транскрипции впереди, после или внутри гена, связывать комплексы белков с метаболитами или гормонами и влиять на конформацию гена. Индукторами или корепрессорами, стимулирующими присоединение регуляторных белков к ДНК, могут быть гормоны, ионы металлов, субстраты или продукты метаболических путей. Определенное значение в регуляции состава и содержания белков имеют посттранскрипционные превращения пре-мРНК в процессе альтернативного сплайсинга, изменение стабильности РНК в разные периоды жизни клетки. Описаны примеры влияния факторов среды на сродство рибосом к мРНК, посттрансляционные модификации полипептидных цепей и изменения продолжительности жизни белковых молекул.
|
|||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 393; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.220.196.68 (0.009 с.) |

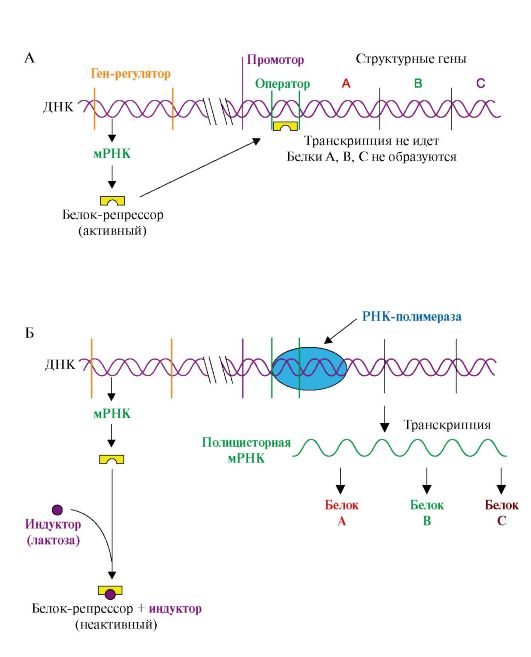 Рис. 3.19. Оперон, регулируемый по механизму индукции (лактозный оперон):
Рис. 3.19. Оперон, регулируемый по механизму индукции (лактозный оперон): Рис. 3.20. Оперон, регулируемый по механизму репрессии:
Рис. 3.20. Оперон, регулируемый по механизму репрессии: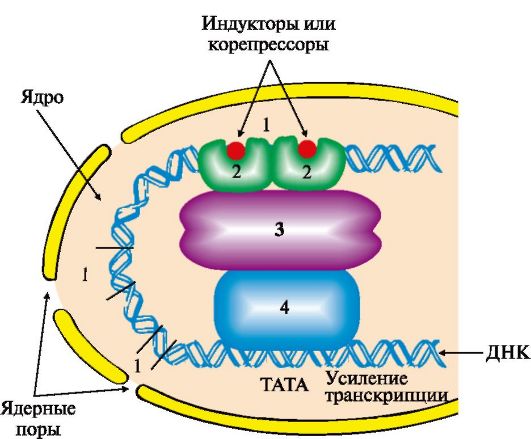 Рис. 3.21. Адаптивная регуляция транскрипции генов у эукариотов.
Рис. 3.21. Адаптивная регуляция транскрипции генов у эукариотов.


