Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Молекулярные механизмы генных, хромосомных и геномных мутацийСодержание книги
Поиск на нашем сайте
Молекулярные механизмы генных, хромосомных и геномных мутаций - раздел Образование, ПАВЛИЧЕНКО В. И АБРАМОВ А. В. Мутационная Изменчивость У Человека. Молекулярные Механизм...
Мутационная изменчивость у человека. Молекулярные механизмы генных мутаций. Классификация генных мутаций. Понятие о моногенных наследственных заболеваниях. Молекулярные и цитологические механизмы хромосомных мутаций. Современные методы изучения кариотипа человека: дифференцированное окрашивание, FISH-метод и др. Классификация мутаций по причинам возникновения. Мутагенные факторы, методы определения мутагенной активности веществ. Антимутагенез. Генеративные и соматические мутации.
Мутационная изменчивость у человека. Стабильность генетического аппарата и обусловливаемый этим аппаратом консерватизм наследственности — лишь одна сторона наследственности. Другая её сторона, столь же неотъемлемая от живого, как и первая, — изменчивость. В совокупности наследственность и изменчивость обеспечили и сохранение жизни на Земле, и непрекращающуюся биологическую эволюцию. Наследственная изменчивость организма обеспечивает необходимую ему приспособляемость к условиям существования как в пределах жизни одного индивида, так и в рамках существования биологического вида в целом. Наследственное многообразие человека — результат длительной эволюции живой материи. При этом надо иметь в виду особенности эволюции человека как биологического и социального существа. У человека как социального существа естественный отбор со временем протекал всё в более специфических формах, что, безусловно, расширяло наследственное разнообразие популяций. Сохранялось то, что могло «отметаться» у животных, или, наоборот, терялось то, что нужно животным. Например, более полноценное обеспечение себя пищей и возможность удовлетворять потребность в витамине С позволили человеку в процессе эволюции «утерять» ген L-гулонолактоноксидазы, катализирующей у животных синтез аскорбиновой кислоты. Наличие этого гена у животных страхует их от развития цинги, а человек из-за такой «всеобщей врождённой ошибки метаболизма» подвержен авитаминозу С. В процессе эволюции человек «приобретал» и нежелательные признаки, имеющие прямое отношение к патологии человека. Большинство видов животных невосприимчиво к дифтерийному токсину и вирусу полиомиелита, потому что у животных отсутствуют компоненты мембраны клеток, обеспечивающие восприятие того или другого патогенного фактора. У человека эти компоненты есть. Гены, их детерминирующие, уже идентифицированы. Например, для восприятия дифтерийного токсина такой ген локализован в 5-й, для вируса полиомиелита — в 19-й хромосоме.
Большинство мутаций увеличивает полиморфизм человеческих популяций (группы крови, цвет волос, рост, разрез глаз и др.), но иногда мутации затрагивают жизненно важные функции, а это уже приводит к болезни. Таким образом, наследственная патология — часть наследственной изменчивости, накопившейся за время эволюции человека. Человек, став биологическим видом Homo sapiens (Человек разумный), как бы заплатил за «сапиентацию» своего вида накоплением патологических мутаций. На основе этих положений формулируется одна из главных концепций медицинской генетики об эволюционном накоплении патологических мутаций в человеческих популяциях. Подтверждением этой концепции являются патологические мутации у животных, по своим проявлениям сходные с наследственными болезнями у человека (ахондроплазии. гемофилии, мышечные дистрофии и др.), а также наличие наследственных болезней у людей, живших несколько тысячелетий назад (о чём можно судить по находкам патологических скелетов в раскопках и произведениям искусства). Эволюция любого вида, в том числе и человека, в конечном счете, сводится к эволюции генотипа. В биологической эволюции человека болезнь как фактор естественного отбора могла играть существенную роль, а эволюция генотипа в свою очередь меняла нозологию патологических процессов. Зависимость эволюции болезни от эволюции генотипа вряд ли может вызывать сомнение. Выше были приведены конкретные формы этой зависимости (цинга, дифтерия, полиомиелит). Факторы эволюции в течение длительного времени влияли не только на формирование биохимических, иммунологических, физиологических или морфологических свойств организма, но и на его патологические реакции, обусловливая значительно большее многообразие нозологических форм болезней у человека по сравнению с таковыми у животных.
Основным источником многообразия наследственных признаков и их непрекращающейся эволюции служит мутационная изменчивость. Способность ДНК мутировать сложилась в эволюции и закрепилась отбором, по-видимому, так же, как и способность противостоять мутационным изменениям, т.е. репарировать их. В организации ДНК заложена возможность ошибок её репликации наряду с возможностью изменения первичной структуры. Вероятность «сбоя» в точности репликации молекулы ДНК невелика: она составляет 10-5—10-7. Однако, принимая во внимание исключительно большое число нуклеотидов в геноме (3,2 х 109 на гаплоидный набор), следует признать, что в сумме на геном клетки на одно её поколение приходится несколько мутаций в структурных генах. По мнению разных авторов, каждый индивид наследует 2—3 новые вредные мутации, которые могут давать летальный эффект или подхватываться отбором, увеличивая генетическое разнообразие человеческих популяций. Изменение нуклеотидной последовательности молекулы ДНК может отразиться на первичной (аминокислотной) структуре белка или на регуляции его синтеза. Так, большой опыт изучения молекулярной природы мутаций гемоглобина показывает, что значительная часть таких мутаций не изменяет функции гемоглобина. Такие мутации нейтральны и не подвергаются отбору. Другие мутации приводят к функциональным отклонениям в молекуле белка. Эти отклонения в каких-то условиях жизни организма могут оказаться полезными, т.е. иметь адаптивное значение, поэтому сохранятся, а иногда и умножатся в последующих поколениях. Именно таким путём возникали и сохранялись в популяциях разнообразные варианты структурных, транспортных и ферментных белков организма. Свойственный организму человека широкий белковый полиморфизм, благодаря которому каждый индивид биохимически неповторим, обусловлен исходно мутационной изменчивостью и отбором адаптивных белковых вариантов. Однако, если структурные отклонения несовместимы с выполнением белком его функции, а она жизненно важна для клетки (организма), мутация становится патологической и в дальнейшем либо исключается из популяции вместе с нежизнеспособной клеткой (организмом), либо сохраняется, обусловливая наследственную болезнь. В отдельных случаях гетерозиготные носители патологической мутации подвергаются положительному отбору. Примером этого служит ген серповидно-клеточной анемии, который широко распространился в популяциях, проживающих в эндемичных по малярии районах, вследствие большой устойчивости гетерозиготных носителей «аномального» гена (мутантного аллеля) к малярийному плазмодию. Различные признаки организма по-разному устойчивы к мутационным изменениям, что связано, по-видимому, со значением признака и с его эволюционным «возрастом». Такие признаки, как гистоновые белки, входящие в состав хромосом, или сократительные белки актин и тубулин, или ферментные белки репликации и транскрипции, весьма консервативны и одинаковы не только у разных представителей человечества, но и у биологических видов значительной филогенетической отдалённости. По-видимому, мутации в соответствующих генах детальны. Большинство белков организма, особенно ферментных, существуют в нескольких изоформах и подвержены таким мутационным изменениям, которые ведут к патологии.
Патологические мутации различны по способности сохраняться и распространяться в популяциях. Одни из них, позволяющие их носителю сохранять плодовитость и не вызывающие серьёзных неблагоприятных сдвигов в фенотипе, могут передаваться из поколения в поколение длительное время. Такие признаки сегрегируют (распределяются) в поколениях согласно законам Менделя, и обусловленный ими генетический груз в популяциях может долго сохраняться. Некоторые комбинации условно патологических рецессивных аллелей могут давать селективное преимущество индивидам (выживаемость, плодовитость). Частота таких аллелей в популяции будет повышаться до определённого уровня в ряду поколений, пока не наступит равновесие между интенсивностью мутационного процесса и отбора. Частота разных мутантных аллелей этого рода может быть неодинаковой в различных популяциях, что определяется популяционными закономерностями (эффект родоначальника, частота кровнородственных браков, миграция и экологические условия). Под эффектом родоначальника подразумевают накопление патологических мутаций в ограниченной популяции от одного носителя болезни группе потомков. Если вновь возникшая мутация имеет доминантное патологическое проявление и ведёт к летальному генетическому исходу (индивид не оставляет потомства), то такой мутационный груз не передаётся следующему поколению. Это обычно доминантные формы тяжёлых болезней, а также большая часть хромосомных болезней. В целом эффекты генетического груза у человека выражены в эволюционно- генетических явлениях сбалансированного полиморфизма, летальности и сниженной фертильности. На основе постоянно протекающих процессов изменения наследственности (мутаций) и отбора генотипов при длительной эволюции человека в популяциях сформировался сбалансированный полиморфизм. Под этим названием понимают такое явление, когда в популяции представлены две формы аллелей одного гена или более, причём частота редкого аллеля составляет не менее 1%. Поскольку возникновение мутаций — редкое событие (1 х 10-7), то, следовательно, частоту мутантного аллеля в популяции более 1% можно объяснить только каким-то селективным преимуществом этого аллеля для организма и постепенным накоплением в ряду поколений после его появления. Примерами сбалансированного полиморфизма являются группы крови АВО, резус, гены муковисцидоза, фенилкетонурии, первичного гемохроматоза. Генетическое многообразие человека основано на сбалансированном полиморфизме, формировавшемся в течение десятков и сотен тысячелетий. Такое многообразие — основа развития человека как биологического вида. Вероятность возникновения и фиксации в популяциях какой-либо мутации с положительным эффектом в эволюционно «отлаженном» человеческом организме существует и в настоящее время, но она крайне мала. Практически новые мутации всегда дают отрицательный эффект.
К эффектам мутационного груза относится летальность. Она проявляется гибелью гамет, зигот, эмбрионов, плодов, смертью детей. Наиболее интенсивно летальные эффекты выражены в человеческих популяциях на уровне зигот. Примерно 60% зигот погибает до имплантации, т.е. до клинически регистрируемой беременности. Исходы всех клинически зарегистрированных беременностей распределяются следующим образом: спонтанные аборты — 15%, мертворождения — 1%, живорождения — 84%. Из 1000 живорождённых детей не менее 5 умирают в возрасте до года по причине наследственной патологии, несовместимой с жизнью. Таков объём летального груза мутационной изменчивости в популяциях человека с медицинской точки зрения. Для большинства наследственных болезней характерна сниженная фертильность, обусловленная нарушением репродуктивной функции. Это ведёт к уменьшенному воспроизводству потомства (и больного, и здорового) в семьях с наследственной патологией. Медицинские и социальные последствия мутационного процесса: социальная дизадаптация (инвалидность) больных, повышенная потребность в медицинской помощи и сниженная продолжительность жизни. Молекулярные механизмы генных мутаций. Генетический материал — ДНК — очень лабилен. Он может меняться, мутировать в результате как внешних, так и внутренних воздействий. Итогом возникающих изменений, если они происходят в соматических клетках (а они происходят непрерывно с самой первой минуты существования нового человеческого организма — зиготы — до последней минуты его жизни), являются многочисленные болезни, включая раковые опухоли и, по-видимому, старение и смерть. Если же они происходят в клетках полового пути, то возникают мутации, которые могут в процессе эволюции закрепляться и распространяться в популяции и приводить к полиморфизму, если они не отсеиваются в силу случайных причин или в силу их вредного воздействия на жизнеспособность индивидуума и его потомства. В целом разнообразие генов зависит от скорости мутаций, размера и демографической истории популяции, в которой происходят мутации, времени, в течение которого происходит накопление этих различий и селекции. Степень разнообразия, которое может поддерживаться в популяции, прямо пропорциональна ее размеру. Сравнительно небольшая вариабельность в популяции человека (вариабельность генома шимпанзе — нашего ближайшего родственника — значительно выше, чем у человека) является результатом ее молодого возраста и происхождения от сравнительно небольшой начальной популяции.
Вредные мутации постоянно возникают, но быстро отсеиваются из популяции. Существует баланс между вновь возникающими мутациями и их отсеиванием селекцией. В результате вредные мутации, вызывающие болезнь, обладают двумя свойствами: они встречаются редко, и каждая конкретная мутация, существующая в популяции, возникла недавно. Что касается обычного полиморфизма, то механизм его поддержания в популяции, несмотря на длительную и интенсивную дискуссию по этому поводу, неясен и, возможно, прояснится, когда удастся достаточно быстро и сравнительно недорого сравнивать множество геномов и провести корреляции между частотами определенных аллелей и историями различных популяций. В молекулах ДНК могут происходить изменения последовательности нуклеотидов. Такие изменения, если они затрагивают функционально активные гены, могут приводить к нарушениям метаболизма или функций (признаков). Если эти изменения не приводят к гибели организма или клетки — они могут передаваться по наследству. Следовательно, генные мутации — это стабильные изменения структуры генов, повторяющиеся в последующих циклах репликации и проявляющиеся у потомства в виде новых вариантов признаков. Все разновидности мутаций связаны с изменением нуклеотидной последовательности генов. Классификация генных мутаций. По особенностям структурных изменений можно отметить несколько групп разнообразных мутаций: · замена одних азотистых оснований другими (транспозиция) (рис.33.); · изменение количества нуклеотидных пар в структуре гена (дупликация, инсерция, делеция); · изменение порядка последовательности нуклеотидов в составе гена (инверсии); · разрыв цепей; · образование сшивок.
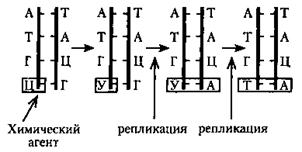
Замена азотистых оснований. Причинами этого рода мутаций являются: а) ошибки репликации, б) влияние определенных химических агентов. Под воздействием химических агентов может происходить нарушение структуры азотистого основания уже присоединенного нуклеотида. Например, под воздействием азотистой кислоты может происходить самопроизвольное дезаминирование цитозина. В результате этого цитозин превращается в урацил. В дальнейшем в цикле репликации урацил соединяется аденином, который в следующем цикле присоединяет тимидиновый нуклеотид. Еще одной причиной может быть ошибочное включение в образующуюся цепь ДНК нуклеотида с измененным основанием. Если это остается незамеченным ферментами репарации, измененное основание включается в процесс репликации, что может привести к замене основной пары на другую.
Рис.33. Схема возникновения мутации (транспозиции) по механизму замены одного азотистого основания другим
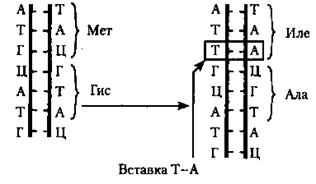
Мутации в результате замены азотистых оснований возникают первоначально в одной из цепей ДНК. Если они не исправляются в ходе репарации, то при последующих репликациях они закрепляются в обеих цепях молекулы. Следствием этого является образование нового триплета в генетическом коде ДНК. Это может отразиться на первичной структуре кодируемого белка, его пространственной организации и функции. Изменения первичной структуры пептида не произойдет в том случае, если новый триплет является «синонимом» прежнего, т. е. будет кодировать ту же аминокислоту. Например, аминокислота лейцин кодируется шестью триплетами: УУА, УУГ, ЦУУ, ЦУЦ, ЦУА, ЦУГ. Замена одного из нуклеотидов в этих триплетах не изменит его «смысла». Этот пример демонстрирует биологическое значение избыточности генетического кода. Однако в большинстве случаев замена одной аминокислоты на другую приводит к серьезным последствиям. Например, замена глутаминовой кислоты валином в молекуле гемоглобина приводит к изменению его структуры и функций. В результате этого у человека развивается болезнь — серповидно-клеточная анемия. В ряде случаев замена азотистых оснований может приводить к появлению нонсенс-кодонов некодирующих аминокислот. Следствием этого будет досрочное прерывание процесса синтеза. Считается, что замена азотистых оснований приводят в - 25 % случаев к образованию триплетов-синонимов, в ~ 5 % случаев — к образованию нонсенс- кодонов, и в ~ 70 % — к возникновению генных мутаций. Изменение количества нуклеотидов в гене. Эта мутация — результат выпадения (делеции) или вставки (инсерции) одной или нескольких пар нуклеотидов в молекулу ДНК (рис. 34). Такой тип мутаций встречается довольно часто. Указанное изменение происходит вследствие воздействия на ДНК некоторых химических агентов, а также радиоактивного облучения. Результатом этой мутации является сдвиг рамки считывания информации с генетического кода. Следствием этого является синтез полипептидов с измененной аминокислотной последовательностью, нарушение структуры и функций белков, нарушение фенотипа. Однако если количество восстановленных или утраченных нуклеотидов кратно трем, то сдвиг рамки не происходит. В этом случае в белке может появиться лишняя аминокислота или будет на одну меньше. Одной из причин мутаций, приводящих к изменению количества нуклеотидов, являются вставки или делеции в результате активности подвижных генетических элементов. Это определенные нуклеотидные последовательности, встроенные в геномы многих организмов. Эти структуры ДНК способны самопроизвольно менять свое положение в результате ошибок при рекомбинации.
Рис.34. Схема изменения количества нуклеотидов
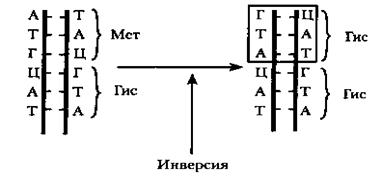
Изменение нуклеотидной последовательности гена (инверсия). Этот тип мутации связан с поворотом определенного участка ДНК на 180°. Такие нарушения происходят вследствие действия химических агентов и ряда физических факторов на молекулярно-генетические Разрывы одной из цепей могут происходить под действием ионизирующей радиации, в результате повреждения химических связей между нуклеотидами (рис. 35). Дефекты могут восстанавливаться ферментом лигазой.
Рис.35. Схема возникновения мутации в результате инверсии
Сшивка нуклеотидов, например, двух рядом стоящих тиминов, происходит под действием ультрафиолетового облучения. Это приводит ошибкам транскрипции. «Мутон». Этим термином называют минимальное количество генетического материала, способного к мутации, что приводит к появлению нового варианта признака. В вышеизложенном материале приведены шовные типы мутаций. Во всех случаях видно, что достаточно изменить только одну пару комплементарных оснований в гене, чтобы ценить свойства белка. То есть мутон соответствует одной паре комплементарных нуклеотидов. Рекон. Этим термином называют минимальное количество генетического материала, изменение которого в результате рекомбинации кроссинговера приводит к мутации и появлению нового варианта признака. Такие процессы могут приводить к сдвигу рамки считывания нарушению синтеза необходимого белка. Научные исследования побывают, что достаточно рекомбинации одной пары комплементарных нуклеотидов, чтобы произошла мутация. Следовательно, рекон соответствует одной паре комплементарных нуклеотидов. Множественные аллели. Различные структурно- фунциональные варианты гена называют аллелями. Они отличаются небольшими изменениями в нуклеотидной последовательности. Это обеспечивает вариации в проявлении признака. Аллели располагаются в одних и тех в участках (локусах) гомологичных хромосом. Наличие в генофонде популяции более двух вариантов аллельных генов называют множественными аллелями. Причиной множественного аллелизма являются разнообразные мутации и рекомбинации. Мутации могут происходить в любых участках гена. Они приводят к тому, что один и тот же ген может участвовать в нескольких вариантах. Если мутации не вызывают гибель организма, они сохраняются в генофонде вида, чем обусловливают появление нового варианта признака в популяции.
|
|||||||||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 132; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.81.79 (0.017 с.) |