
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Фосфолипиды, строение, биологическая роль.
Фосфолипиды - молекулы, обладающие амфифильными свойствами, так как они имеют гидрофобную часть, образованную чаще всего радикалами жирных кислот, и гидрофильную часть - остаток фосфорной кислоты, аминоспирта или аминокислоты. Фосфолипиды могут вследствие своих амфифильных свойств образовывать бислойные структуры мембран или гидрофильный монослой на поверхности липопротеинов - частиц, обеспечивающих транспорт гидрофобных липидов кровью. Фосфолипиды разделяют на глицерофосфолипиды, основой строения которых является глицерол, и сфинголипиды, имеющие в своей структуре аминоспирт сфингозин (табл. 8.1). Сфинголипиды содержатся в мембранах многих клеток; в наружных мембранах они являются составной частью антигенов Таблица 8.1. Строение и функции основных классов липидов человека
и рецепторов. Особенно много сфинголипидов в нервной ткани, где они формируют миелиновые оболочки нейронов. Холестерол - основной стероид и предшественник всех остальных стероидов в организме человека. Холестерол входит в состав мембран всех клеток, влияя на свойства их гидрофобного слоя, и является предшественником в синтезе желчных кислот, стероидных гормонов. Биосинтез и использование кетоновых тел. Изменение их концентрации в крови и моче при патологии. КЕТОАЦИДОЗ 1. К кетоновым телам относят три вещества: β-гидроксибутират, ацетоацетат и ацетон.
Только первые два являются источниками энергии и могут окисляться в тканях. В норме концентрация кетоновых тел в крови невелика и составляет 1-3 мг/дл. Синтез кетоновых тел увеличивается при: • голодании; • длительной интенсивной физической нагрузке; • употреблении пищи, богатой жирами, но с низким содержанием углеводов (диета Аткинса, «кремлевская» диета); • сахарном диабете. Кетоновые тела являются водорастворимыми кислотами, поэтому в отличие от жирных кислот, они могут проходить через гематоэнцефалический барьер и служат, наряду с глюкозой, источником энергии для нервной ткани, особенно после 3-5 дней голодания, когда концентрация кетоновых тел в крови существенно увеличивается (рис. 8.20). Скелетные мышцы и почки используют кетоновые тела даже при их низкой концентрации в крови.
2. Синтез кетоновых тел происходит в митохондриях печени и в небольшом количестве - в корковом слое почек. Исходным субстратом синтеза служит ацетил-КоА, образующийся в результате β-окисления жирных кислот. Ситуация, в которой увеличивается синтез кетоновых тел, развивается следующим образом. При голодании гормон глюкагон (или адреналин при физической работе) через аденилатциклазную систему в жировой ткани активирует распад жира. Жирные кислоты выделяются в кровь и транспортируются в комплексе с альбуминами в печень. В печени увеличивается скорость β-окисления и образуется большое количество ацетил-КоА. Скорость реакций цикла Кребса в этих условиях снижена в результате ингибирования регуляторных ферментов цитратного цикла аллостерическими ингибиторами АТФ и NADH, концентрация которых повышена в результате активного β-окисления. Кроме того, при высокой концентрации NADH оксалоацетат восстанавливается до малата и в такой форме переносится в цитозоль, где реакция идет в обратном направлении. Оксалоацетат становится субстратом для глюконеогенеза и менее доступен для взаимодействия с ацетил-КоА. В результате в митохондриях накапливается ацетил-КоА, который используется для синтеза кетоновых тел (рис. 8.21, 8.22).
Пунктирная линия - скорость метаболических путей снижена, сплошная линия - повышена. При голодании в результате преобладания действия глюкагона активируются липолиз в жировой ткани и β-окисление в печени. Количество оксалоацетата в митохондриях уменьшается, так как его образуется меньше, и, кроме того, он выходит в цитозоль (восстанавливаясь до малата), где используется в глюконеогенезе. В результате скорость использования ацетил-КоА в ЦТК снижается и он используется для синтеза кетоновых тел. Синтез кетоновых тел значительно увеличивается при сахарном диабете Основным кетоновым телом в крови является β-гидроксибутират (рис. 8.22), так как равновесие в реакции Ацетоацетат β-Гидроксибутират сдвинуто вправо из-за присутствия высоких концентраций NАDН, который восстанавливается в реакциях β-окисления, протекающих, как и синтез кетоновых тел, в матриксе митохондрий. Повышение концентрации кетоновых тел в крови называют кетонемией. 3. Регуляция синтеза кетоновых тел осуществляется через фермент ГМГ-КоА-синтазу (рис. 8.22). Этот фермент ингибируется при высоких концентрациях свободного кофермента HS-КоА. Таким образом скорость синтеза кетоновых тел координируется с количеством жирных кислот, поступающих в печень. В ситуациях голодания, сахарного диабета, тяжелой физической работы под действием гормонов глюкагона и адреналина происходит мобилизация ТАГ из жировой ткани, поток жирных кислот в печень увеличивается и HS-КоА связывается с жирными кислотами в ацилКоА. Концентрация свободного HS-КоА снижается и синтез кетоновых тел увеличивается.
* - реакция происходит неферментативно только при высокой концентрации ацетоацетата в крови, например при длительном голодании или сахарном диабете 4. Окисление кетоновых тел как источников энергии происходит во многих тканях (рис. 8.23). В печени отсутствует фермент, необходимый для активации кетоновых тел - сукцинил-КоА-ацетоацетаттрансфераза. Поэтому печень не окисляет кетоновые тела и продуцирует их только на «экспорт». Эритроциты, в которых отсутствуют митохондрии, также не используют кетоновые тела. 5. При длительном голодании и особенно при сахарном диабете в крови существенно возрастает концентрация кетоновых тел и организм не успевает их утилизировать. При накоплении кетоновых тел развивается кетоацидоз, так как ацетоацетат и β-гидроксибутират - это легко диссоцирующие кислоты:
Выведение кетоновых тел, в том числе и ацетона, с мочой (кетонурия), потом выдыхаемым воздухом является способом выведения избытка кетоновых тел из организма и уменьшения таким образом ацидоза.
При окислении кетоновые тела активируются путем превращения ацетоацетата в ацетоацетил-КоА. Донором КоА является сукцинил-КоА. В результате окисления β-гидроксибутирата образуется 2 ацетил~КоА, которые далее окисляются в ЦТК. Таким образом при окислении β-гидроксибутирата образуется 27 молекул АТФ, но для активации ацетоацетата используется энергия одной макроэргической связи сукцинил-КоА, поэтому теоретический выход АТФ составляет 26 молекул АТФ
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 124; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.117.158.47 (0.007 с.) |
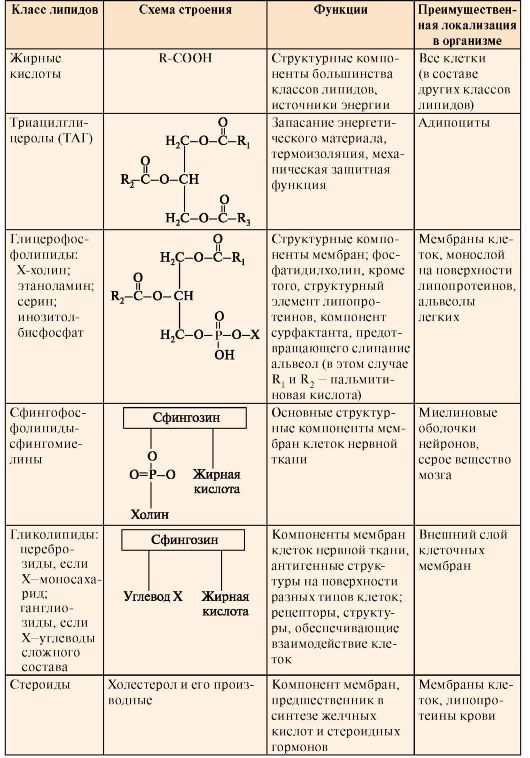 Таблица 8.2. Состав и строение жирных кислот организма человека
Таблица 8.2. Состав и строение жирных кислот организма человека Примечание. Сп - число атомов углерода в данной жирной кислоте; m - число двойных связей в радикале жирной кислоты; Δ9, 12 - положение двойных связей в радикале жирной кислоты, считая от первого, карбоксильного углерода; ω-3, ω-6 - положение первой двойной связи, считая от метильного углерода радикала жирной кислоты; знаком * отмечены полиеновые эссенциальные жирные кислоты, которые должны поступать с пищей. Остальные полиеновые кислоты могут синтезироваться из них, например, арахидоновая кислота может синтезироваться из линолевой (18:2 ω-6). В колонке ** представлен суммарный состав жирных кислот у человека, находящегося на обычном пищевом рационе. В зависимости от типа ткани, жирных кислот пищи и ряда других условий состав жирных кислот липидов может несколько отличаться от указанного
Примечание. Сп - число атомов углерода в данной жирной кислоте; m - число двойных связей в радикале жирной кислоты; Δ9, 12 - положение двойных связей в радикале жирной кислоты, считая от первого, карбоксильного углерода; ω-3, ω-6 - положение первой двойной связи, считая от метильного углерода радикала жирной кислоты; знаком * отмечены полиеновые эссенциальные жирные кислоты, которые должны поступать с пищей. Остальные полиеновые кислоты могут синтезироваться из них, например, арахидоновая кислота может синтезироваться из линолевой (18:2 ω-6). В колонке ** представлен суммарный состав жирных кислот у человека, находящегося на обычном пищевом рационе. В зависимости от типа ткани, жирных кислот пищи и ряда других условий состав жирных кислот липидов может несколько отличаться от указанного Рис. 8.20. Изменение концентрации глюкозы, жирных кислот и кетоновых тел в плазме крови при голодании
Рис. 8.20. Изменение концентрации глюкозы, жирных кислот и кетоновых тел в плазме крови при голодании Рис. 8.21. Активация синтеза кетоновых тел при голодании
Рис. 8.21. Активация синтеза кетоновых тел при голодании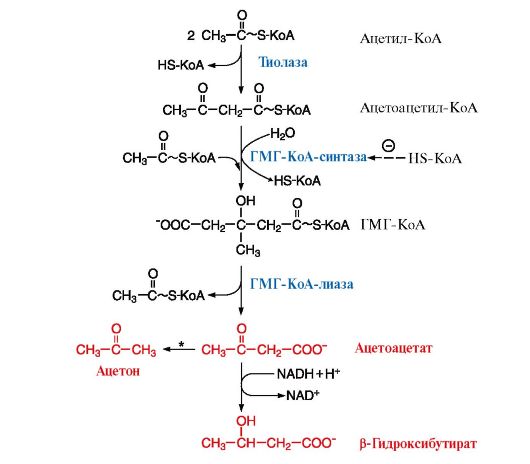 Рис. 8.22. Синтез кетоновых тел в митохондриях гепатоцитов
Рис. 8.22. Синтез кетоновых тел в митохондриях гепатоцитов Кетоацидоз является опасным осложнением сахарного диабета. При высоких концентрациях кетоновых тел в крови ацетоацетат неферментативно декарбоксилируется, превращаясь в третье кетоновое тело - ацетон. Ацетон не утилизируется как источник энергии и выводится из организма.
Кетоацидоз является опасным осложнением сахарного диабета. При высоких концентрациях кетоновых тел в крови ацетоацетат неферментативно декарбоксилируется, превращаясь в третье кетоновое тело - ацетон. Ацетон не утилизируется как источник энергии и выводится из организма. Рис. 8.23. Схема окисления кетоновых тел
Рис. 8.23. Схема окисления кетоновых тел


