
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Биосинтез жиров в печени и жировой ткани. Регуляция синтеза жиров
1. Жиры - наиболее компактная форма запасания энергетического материала, поэтому избыточное количество углеводов, получаемых с пищей, перерабатывается в жиры и запасается в адипоцитах. Синтез жиров происходит в абсорбтивный период и стимулируется инсулином. Наиболее активно синтез жиров происходит в печени, жировой ткани и лактирующей молочной железе. 2. Для синтеза жиров необходимы глицерол в виде глицерол-3-фосфата (рис. 8.13) и активные формы жирных кислот - ацил-КоА. Глицерол вначале должен превратиться в глицерол-3-фосфат. В печени глицерол-3-фосфат образуется двумя путями: а) при восстановлении дигидроксиацетонфосфата (метаболита гликолиза); б) при фосфорилировании глицеролкиназой свободного глицерола, попадающего в печень из крови. В жировой ткани фермент глицеролкиназа отсутствует и единственным источником глицерол-3-фосфата является дигидроксиацетонфосфат, образующийся в процессе гликолиза, поэтому одновременно с синтезом жиров в адипоцитах должны проходить реакции гликолиза. 3. В печени жирные кислоты, необходимые для синтеза жиров, синтезируются в основном из продуктов катаболизма глюкозы. Далее синтез жиров идет через образование фосфатидной кислоты (рис. 8.13). Синтезированные в гепатоцитах жиры упаковываются в незрелые ЛПОНП, главным белковым компонентом которых является апоВ-100. Этот белок кодируется тем же геном, что и белок В-48 в слизистой тонкой кишки, однако в гепатоцитах он соответствует всей кодирующей части гена (100%), а в кишечнике в результате посттранскрипционных изменений считывается только 48% гена, поэтому этот белок называют апоВ-48. В крови ЛПОНП получают апоЕ и С-II от ЛПВП и превращаются в зрелые ЛПОНП, функцией которых является транспорт кровью жиров, синтезированных в печени, в другие органы, главным образом в жировую ткань. 4. В капиллярах жировой и других тканей жиры в составе ЛПОНП и хиломикронов подвергаются гидролизу под действием липопротеинлипазы. Этот фермент «узнает» на поверхности липопротеинов белок апоС-II, активируется им и гидролизует ТАГ, входящие в состав липопротеинов, до глицерола и жирных кислот. В разных тканях находятся изоформы ЛП-липазы, отличающиеся значениями Кш. В капиллярах жировой ткани значение Кm выше - следовательно, этот фермент активно работает при высокой концентрации липопротеинов в крови в абсортивный период; в мышцах Кш ниже - следовательно, мышцы могут утилизировать жирные кислоты, освобождающиеся из липопротеинов, и при более низкой их концентрации в крови. Жирные кислоты, освободившиеся под действием
Синтез жиров в печени происходит в основном из продуктов катаболизма глюкозы. После еды образующийся в результате гликолиза избыток ацетил-КоА активно используется для синтеза жирных кислот (1). Глицерол-3-фосфат образуется двумя путями (2, 3). Синтез ТАГ происходит через образование фосфатидной кислоты (4), при дефосфорилировании которой образуется ДАГ (5). Следующая реакция ацилирования превращает его в ТАГ (6). ТАГ в составе ЛПОНП секретируются в кровь ЛП-липазы, из крови поступают в клетки и используются по-разному: в адипоцитах - для синтеза жиров и их депонирования (рис. 8.14), а в миокарде и скелетных мышцах они окисляются, образуя АТФ, необходимый для работы этих тканей. 5. Запасание жиров в жировой ткани - так называемое депонирование жиров - происходит в абсорбтивный период, когда увеличивается соотношение инсулин - глюкагон. Инсулин активирует: • синтез ЛП-липазы и ее экспонирование на поверхности капилляров в жировой ткани; • транспорт глюкозы внутрь адипоцитов, стимулируя включение ГЛЮТ-4 в плазматическую мембрану, так как жировая ткань является инсулинзависимой; • гликолиз, что обеспечивает образование глицерол-3-фосфата и АТФ, необходимых для синтеза жиров; • пентозофосфатный путь, в ходе которого восстанавливается NADP+, необходимый для синтеза жирных кислот. 6. Сравнение гликогена и жиров как запасаемых энергоносителей. По существу роль гликогена и жиров в организме одинакова - это формы запасания энергии. Однако между ними есть и значительные различия как в количественном, так и в функциональном отношении.
Таблица 8.5. Гликоген и жиры в организме человека
Если учесть, что жиры и по калорийности превосходят углеводы, то разница в запасе энергии в этих формах становится еще внушительнее. Запаса гликогена хватает примерно на 1 сутки голодания, в то время как жиров - на много недель.
Суточное потребление углеводов превышает содержание гликогена в организме. Это значит, что полное обновление гликогена в организме может произойти менее чем за 2 суток (например, после суточного голодания и последующих приемов пищи в течение дня). За 1 сутки может обновиться только 1/100 всего запаса жиров. Запасы гликогена в клетках расходуются на всем протяжении суток, за исключением примерно двухчасовых промежутков после приемов пищи. Жиры, депонированные в жировой ткани, могут и не расходоваться, поскольку при обычном ритме питания в крови постоянно имеются липопротеины, снабжающие органы жирными кислотами. Таким образом, можно считать, что липопротеины выполняют не только транспортную функцию, но и функцию краткосрочного запасания жиров. По роли в энергетическом обмене жиры липопротеинов (хиломикронов и ЛПОНП) в большей мере сходны с гликогеном, чем жиры, депонированные в жировой ткани. Важной особенностью жиров является то, что при их гидролизе образуется два функционально различных продукта - жирные кислоты и глицерол. Глицерол используется для глюконеогенеза (наряду с аминокислотами) и тем самым участвует в обеспечении глюкозой клеток мозга и других глюкозозависимых клеток при голодании, а окисление жирных кислот является источником АТФ для многих тканей. 59.Транспортные липопротеины крови; особенности состава и функции Роль аполипопротеинов и липопротеинлипазы. для транспорта липидов кровью в организме образуются комплексы липидов с белками - липопротеины. Все типы липопротеинов имеют сходное строение - гидрофобноеядро и гидрофильныйслой на поверхности. В организме синтезируются следующие типы липопротеинов: хиломикроны (ХМ), липопротенны очень низкой плотности (ЛПОНП), липопротеины промежуточной плотности (ЛППП), липопротеины низкой плотности (ЛПНП) и липопротеины высокой плотности (ЛПВП). Каждый из типов ЛП образуется в разных тканях и транспортирует определённые липиды. Например, ХМ транспортируют экзогенные (пищевые жиры) из кишечника в ткани, поэтому триацнлглицеролы составляют до 85% массы этих частиц. ЛП хорошо растворимы в крови, не коалесцируют, так как имеют небольшой размер и отрицательный заряд на поверхности. Некоторые ЛП легко проходят через стенки капилляров кровеносных сосудов и доставляют липиды к клеткам. Большой размер ХМ не позволяет им проникать через стенки капилляров, поэтому из клеток кишечника они сначала попадают в лимфатическую систему и потом через главный грудной проток вливаются в кровь вместе с лимфой. Гидрофильный слой образован белками, которые называют апопротеинами, и амфифильными молекулами липидов - фосфолипидами и холестеролом. Гидрофильные группы этих молекул обращены к водной фазе, а гидрофобные части - к гидрофобному ядру липопротеина, в котором находятся транспортируемые липиды.
Метод ультрацентрифугирования позволяет разделить ЛП, используя их различие в плотности, которая зависит от соотношения количества липидов и белков в частице. Так как жир имеет меньшую, чем вода, плотность, то ХМ, содержащие более 85% жиров, располагаются на поверхности сыворотки крови, а ЛПВП, содержащие наибольшее количество белков, имеют самую большую плотность и при центрифугировании располагаются в нижней части центрифужной пробирки.
метод электрофореза. Скорость движения частиц при электрофорезе зависит от их заряда и размера. Заряд, в свою очередь, зависит от количества белков на поверхности ЛП (табл. 8-5). При электрофорезе в геле все типы ЛП движутся к положительному полюсу; ближе к старту располагаются ХМ, а ЛПВП, имеющие наибольшее количество белков и наименьший размер, удаляются от старта дальше других частиц. Все липопротеины имеют сходное строение: ядро состоит из гидрофобных молекул: ТАГ, эфиров холестерола, а на поверхности находится монослой фосфолипидов, полярные группы которых обращены к воде, а гидрофобные погружены в гидрофобное ядро липопротеина (рис. 8.3). Кроме фосфолипидов, на поверхности находятся белки - аполипопротеины (табл. 8.3). Аполипопротеины выполняют различные функции. Интегральные аполипопротеины являются структурными компонентами. Периферические аполипопротеины в плазме крови могут передаваться от одного типа липопротеинов к другим, определяя их дальнейшие превращения. Таблица 8.3. Характеристика липопротеинов
|
|||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 276; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.127.232 (0.012 с.) |
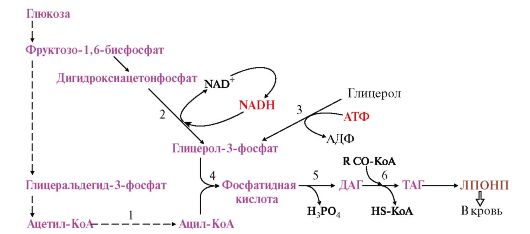 Рис. 8.13. Синтез жиров в печени
Рис. 8.13. Синтез жиров в печени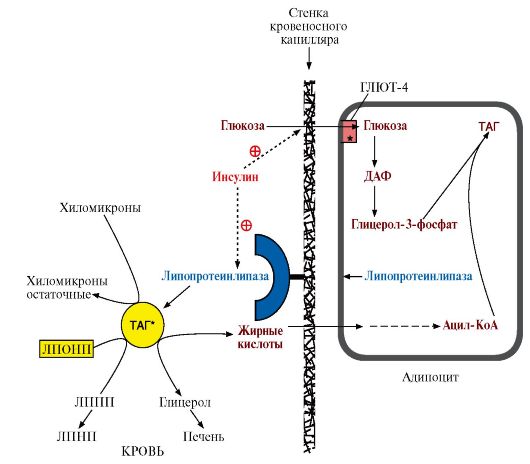
 Жиров в организме содержится в приблизительно в 30 раз больше, чем гликогена (табл. 8.5.)
Жиров в организме содержится в приблизительно в 30 раз больше, чем гликогена (табл. 8.5.)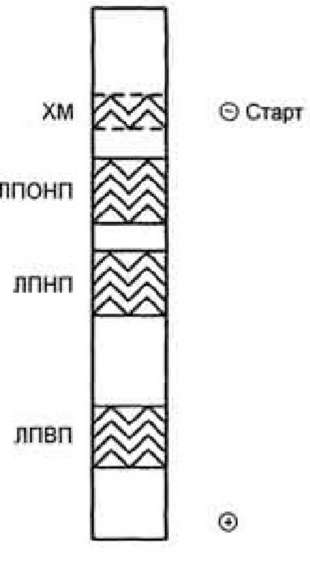 Апопротеины выполняют несколько функций: формируют структуру липопротеинов; взаимодействуют с рецепторами на поверхности клеток и таким образом определяют, какими тканями будет захватываться данный тип липопротеинов; служат ферментами или активаторами ферментов, действующих на липопротеины
Апопротеины выполняют несколько функций: формируют структуру липопротеинов; взаимодействуют с рецепторами на поверхности клеток и таким образом определяют, какими тканями будет захватываться данный тип липопротеинов; служат ферментами или активаторами ферментов, действующих на липопротеины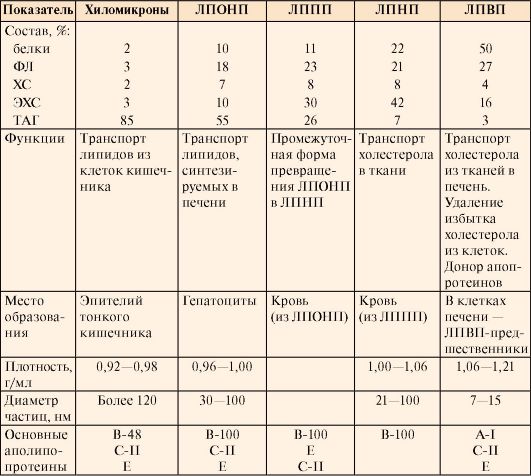
 О
О


