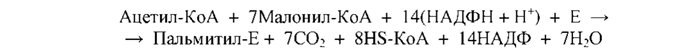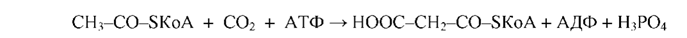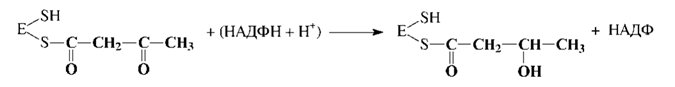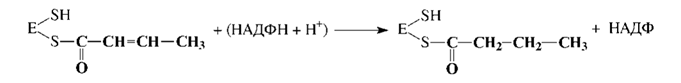Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Окисление жирных кислот с четным числом углеродных атомов.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют четное число углеродных атомов. Любая такая кислота, от которой отщепляется по паре углеродных атомов, в конце концовпроходит через стадию масляной кислоты. После очередного β-окисления масляная кислота становится ацетоуксусной. Последняя затем гидролизуется до двух молекул уксусной кислоты. Окисление жирных кислот с нечетным числом углеродных атомов. Как отмечалось, основная масса природных липидов содержит жирные кислоты с четным числом углеродных атомов. Однако в липидах многих растений и некоторых морских организмов присутствуют жирные кислоты с нечетным числом атомов углерода. Кроме того, у жвачных животных при переваривании углеводов в рубце образуется большое количество пропионовой кислоты, которая содержит три углеродных атома. Пропионатвсасывается в кровь и окисляется в печени и других тканях. Установлено, что жирные кислоты с нечетным числом углеродных атомов окисляютсятаким же образом, как и жирные кислоты с четным числом углеродных атомов, с той лишь разницей, что на последнем этапе расщепления (β-окисления) образуется одна молекула пропионил-КоА и одна молекула ацетил-КоА, а не 2 молекулы ацетил-КоА. Активированный трехуглеродный фрагмент – пропионил-КоА – включается в цикл трикарбоновых кислот после превращения в сукцинил-КоА. Болезни связанные с нарушением окисления жирных кислот. Изучение митохондриальныхболезней, обусловленных нарушением бета окисления жирных кислот с различной длиной углеродной цепи было начато в 1976 г., когда учёные впервые описали больных с дефицитом ацил-КоА-дегидрогеназысреднецепочечных жирных кислот и глутаровойацидемией II типа. В настоящее время эта группа заболеваний включает не менее 12 самостоятельных нозологических форм, происхождение которых связано с генетически детерминированными расстройствами трансмембранного транспорта жирных кислот (системный дефицит карнитина, дефицит карнитинпальмитоилтрансфераз I и II, ацилкарнитин-карнитинтранслоказы) и их последующего митохондриального бета-окисления (дефицит ацил-КоА и 3-гидрокси-ацил-КоА-дегидрогеназ жирных кислот с различной длиной углеродной цепи, глутароваяацидемия II типа). Частота дефицита ацил-КоАдегидрогеназысреднецепочечных жирных кислот составляет 1:8900 новорождённых, частота других форм патологии пока не установлена.
Генетические данные и патогенез. Заболевания имеют аутосомно-рецессивный тип наследования. Патогенез болезней обмена жирных кислот связан с истощением углеводных запасов в условиях метаболического стресса (интеркуррентных инфекционных болезней, физической или эмоциональной перегрузке, голодании, хирургическом вмешательстве). В подобной ситуации липиды становятся необходимым источником восполнения энергетических потребностей организма. Происходит активация дефектных процессов транспорта и бета-окисления жирных кислот. Вследствие мобилизации омега-окисления происходит накопление в биологических жидкостях дикарбоновых кислот, их токсичных производных, конъюгатов карнитина - в результате развивается вторичная карнитиновая недостаточность. Симптомы. Клинические проявления всех болезней обмена жирных кислот имеют большое сходство. Заболевания, как правило, характеризуются приступообразным течением. Существуют тяжёлая (ранняя, генерализованная) и лёгкая (поздняя, мышечная) формы, отличающиеся разной степенью ферментного дефицита или его тканевой локализацией. Тяжёлая форма манифестирует в раннем возрасте, в том числе в периоде новорождённости. Основные симптомы: рвота, генерализованные тонико-клонические судороги или инфантильные спазмы, прогрессирующие вялость, сонливость, общая мышечная гипотония, нарушение сознания вплоть до комы, расстройство сердечной деятельности (нарушение ритма или кардиомиопатия), увеличение печени (синдром Рея). Заболевание сопровождается летальностью (до 20%) и риском внезапной детской смерти. Лёгкая форма обычно впервые проявляется в школьном возрасте и у подростков. Развиваются боли в мышцах, слабость, утомляемость, моторная неловкость, тёмная окраска мочи (миоглобинурия). Характерные дополнительные клинические признаки дефицита 3-гидрокси-ацил-Ко А-дегидрогеназы жирных кислот с длинной углеродной цепью - периферическаянейропатия и пигментный ретинит. У будущих матерей, дети которых, вероятно, будут иметь этот ферментный дефект, нередко осложняется течение беременности - развиваются жировая инфильтрация печени, тромбоцитопения, повышается активность трансаминаз.
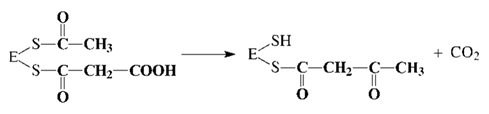
63.биосинтез жирных кислот. Особенности синтеза ненасыщенных жирных кислот. Незаменимые жирные кислоты. Синтез длинноцепочечных насыщенных и ненасыщенных жирных кислот. БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ Биосинтез жирных кислот и липидов играет важную роль в жизнедеятельности организмов. Именно в виде жирных кислот и триацилглицеринов откладываются основные количества энергетических ресурсов организмов животных, в то время как энергоресурсы, откладываемые в форме углеводов, незначительны. В клетках организма жирные кислоты синтезируются из ацетил-КоА, образующегося из избыточной глюкозы пищи, которая не была использована организмом на энергетические нужды. В качестве восстановителя в биосинтезе жирных кислот принимает участие НАДФН, синтезируемый, в основном, в пентофосфатном пути распада углеводов. Нужно отметить, что хотя все реакции β-окисления жирных кислот обратимы, этот путь не используется организмом с целью их синтеза. Биосинтез жирных кислот осуществляется в цитоплазме клеток и катализируется целым полиферментным надмолекулярным ансамблем - пальмитилсинтетазой, состоящей из семи ферментов. Суммарная реакция биосинтеза жирных кислот в цитоплазме имеет следующий вид (Е - пальмитилсинтетаза):
Из данного уравнения можно видеть, что для синтеза жирной кислоты требуется всего одна молекула ацетил-КоА, служащая «затравкой». Непосредственным источником синтеза является малонил-КоА, который образуется из ацетил-КоА по реакции:
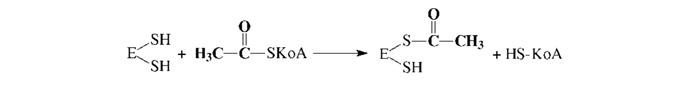
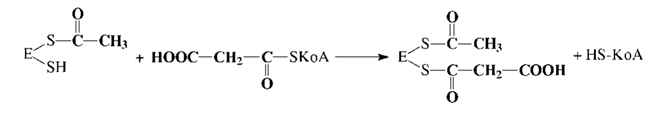
Эта реакция катализируется биотинзависимым ферментом - ацетил-КоА- карбоксилазой. Функция биотина сводится к переносу диоксида углерода на субстрат. Пальмитилсинтетаза представляет собой многофункциональный ансамбль белков: в центре полиферментного ансамбля находится ацилпереносящий белок (АПБ), содержащий свободную SH-группу; шесть остальных ферментов располагаются по периметру, причем один из них также содержит SH- группу. Процесс синтеза жирной кислоты описывается рядом последовательных реакций: 1. Перенос ацетила с ацетил-КоА на синтетазу: 2. Перенос малонила с малонил-КоА на синтетазу: 3. Конденсация ацетила с малонилом и декарбоксилирование образовавшегося продукта:
4. Первое восстановление промежуточного продукта с участием НАДФН:
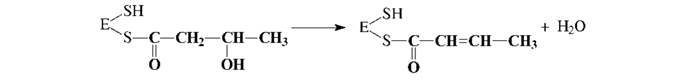
5. Дегидратация промежуточного продукта: 6. Второе восстановление промежуточного продукта с участием НАДФН: Затем синтезированный бутирил перемещается на ту SH-группу, с которой был связан затравочный ацетил, а на освободившуюся SH-группу поступает новый малонильный остаток из малонил-КоА. Далее цикл повторяется снова; после семи оборотов цикла синтезируется пальмитил-Е, который при участии пальмитилдеацилазыгидролизуется до пальмитиновой кислоты и фермента (Е). Пальмитиновая кислота - это основной продукт биосинтеза, однако в небольших количествах могут образовываться и другие жирные кислоты. Жирные кислоты с разветвленной углеродной цепью синтезируются из продуктов метаболизма аминокислот с разветвленной цепью (валин, изолейцин и лейцин) через ацильные производные КоА путем удлинения цепи и при участии АПБ. Особенности биосинтеза полиненасыщенных жирных кислот представляют интерес в связи с их витаминоподобными функциями. Некоторые полиеновые кислоты могут синтезироваться из олеиновой кислоты с помощью ряда последовательных реакций. Однако, синтез полиненасыщенных кислот, содержащих двойные связи, расположенные между конечным метилом и седьмым атомом углерода, невозможен, поэтому они и являются незаменимыми в пищевом рационе. Таким образом, биосинтез и поступление с пищей - два основных источника жирных кислот для организма человека и животных. Образование ненасыщенных жирных кислот. В отличие от растительных тканей ткани животных обладают весьма ограниченной способностью превращать насыщенные жирные кислоты в ненасыщенные. Установлено, что две наиболее распространенные мононенасыщенные жирные кислоты – пальмитоолеиновая и олеиновая – синтезируются из пальмитиновой и стеариновой кислот.Эти превращения протекают в микросомах клеток печени и жировой ткани при участии молекулярного кислорода, восстановленной системы пиридиновых нуклеотидов и цитохрома b5. Превращению подвергаются только активированные формы пальмитиновой и стеариновой кислот. Ферменты, участвующие в этих превращениях, получили название десатураз. Наряду с десатурацией жирных кислот (образование двойных связей) в микросомах происходит и их удлинение (элонгация), причем оба эти процесса могут сочетаться и повторяться. Удлинение цепи жирной кислотыпроисходит путем последовательного присоединения к соответствующемуацил-КоАдвууглеродных фрагментов при участии малонил-КоА и НАДФН. Энзиматическая система, катализирующая удлинение жирных кислот, получила название элонгазы. На схеме представлены пути превращения пальмитиновой кислоты в реакциях десатурации и элонгации. Незаменимые жирные кислоты. В настоящее время показано, что в микросомах клеток млекопитающих образование двойных связей может происходить только на участке цепи жирной кислоты от 9-го до 1-го углеродных атомов, ибо в микросомах отсутствуют десатуразы, которые могли бы катализировать образование двойных связей в цепи далее 9-го углеродного атома. У животных двойные связи могут образовываться в Δ4-, Δ5-, Δ6- и Δ9-положении, но не далее Δ9-положения, в то время как у растений – в Δ6-, Δ9-, Δ12 и Δ15-положении. Поэтому в организме млекопитающих, в том числе и человека, не могут образовываться, например, из стеариновой кислоты (18:0) линолевая (18:2; 9,12) и линоленовая (18:3; 9,12,15) кислоты. Эти кислоты относятся к категории незаменимых жирных кислот. К незаменимым жирным кислотам обычно относят также арахидоновую кислоту (20:4; 5,8,11,14). У большинства млекопитающих арахидоноваякислота может образовываться из линолевой кислоты. Незаменимые жирные кислоты должны поступать в организм с пищей. При длительном их отсутствии в пище у животных наблюдается отставание в росте, развиваются характерные поражения кожи и волосяного покрова. Описаны случаи недостаточности незаменимых жирных кислот и у человека. Так, у детей грудного возраста, получающих искусственное питание с незначительным содержанием жиров, может развиться чешуйчатый дерматит, который поддается лечению препаратом линолевой кислоты. Нарушения, обусловленные недостатком незаменимых жирных кислот, наблюдаются также у больных, жизнедеятельность которых в течение длительного времени поддерживается только за счет внутривенного питания, почти лишенного жирных кислот. Принято считать, что во избежание этих нарушений необходимо, чтобы на долю незаменимых жирных кислот приходилось не менее 1–2% от общей потребности в калориях. Следует отметить, что незаменимые жирные кислоты содержатся в достаточнобольших количествах в растительных маслах. Исследования, проведенные с применением изотопов, показали, что арахидоновая кислота и некоторые другие 20-углеродные (эйкозановые) кислоты, содержащие двойные связи, участвуют в образовании эйкозаноидов.
64. метаболизм сложных липидов. Наследственные болезни, связанные с нарушением катаболизма сложных липидов. МЕТАБОЛИЗМ ФОСФОЛИПИДОВ В отличие от триглицеридов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды играют важную роль в структуре и функции клеточных мембран, активации мембранныхи лизосомальных ферментов, в проведении нервных импульсов, свертывании крови, иммунологических реакциях, процессах клеточной пролиферации и регенерации тканей, в переносе электронов в цепи «дыхательных» ферментов. Особая роль фосфолипидам отводится в формировании липопротеидных комплексов. Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, семенниках, яичниках, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазматической сети клетки. Центральную роль в биосинтезе фосфолипидов играют 1,2-диглицериды (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатиднаякислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфингомиелинов). Цитидинтрифосфат (ЦТФ) участвует в синтезе практически всех фосфолипидов. В качестве примера рассмотрим синтез отдельных представителей фосфолипидов. Гликолипидозы – группа заболеваний, обусловленных нарушением распада гликолипидов (жироподобные вещества, содержащие углеводы).
|
||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 2564; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.93.18 (0.01 с.) |