Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Ветеринарний контроль при штучному осіменінніСодержание книги
Поиск на нашем сайте
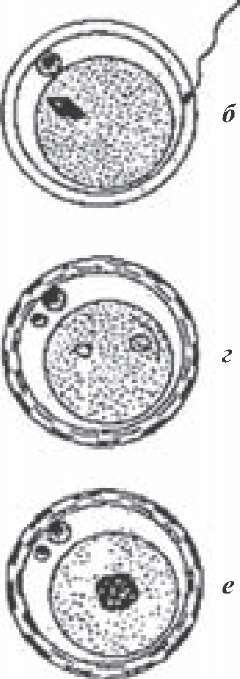
Для охорони здоров'я плідників, попередження занесення на племоб'єднання та розповсюдження через штучне осіменіння заразних захворювань і забезпечення нормального відтворення тварин потрібно суворо дотримуватися наявних ветеринарно- санітарних правил при комплектуванні племпідприємств плідниками, їх утриманні та використанні, одержанні від них сперми, технологічній її обробці та осіменінні тварин. У комплексі виробничих приміщень племпідприємства повинен бути ветеринарний пункт, карантин та ізолятор. Племпідприємство є установою закритого типу. Доступ на його територію сторонніх осіб та транспорту забороняється. Для проходу на племпідприємство обслуговуючого персоналу влаштовують санпропускник, а для в'їзду транспорту - дезбар'єр. Комплектують племпідприємства здоровими тваринами з безпечних щодо заразних захворювань господарств. Перед вивезенням плідників з господарства кожного з них піддають клінічному дослідженню, що включає загальний огляд, дослідження статевих органів, рефлексологічну пробу на самку (статево дозрілих самців) та оцінку сперми. При дослідженні статевих органів звертають увагу на розвиток сім'яників, їх симетрію. Під час пробного статевого акту звертають увагу на прояви статевих рефлексів. Завезених тварин карантинують протягом 30 днів (завезених з-за кордону - 60 днів), досліджуючи їх ще раз на небезпечні хвороби. Протягом карантинування тварин утримають у спеціальних (карантинних) приміщеннях, закріплюють за ними окремий обслуговуючий персонал, а також забезпечують відповідний санітарний режим у приміщенні. Після проведення діагностичних досліджень, при їх негативних наслідках, плідників вакцинують у відповідності з планом протиепізоотичних заходів. Щоб своєчасно помічати всі випадки захворювань плідників, перед кожним використанням потрібно піддавати їх ветеринарному огляду. Всі приміщення, манеж, предмети догляду та інвентар періодично обробляють та дезінфікують, плідників щоденно чистять, а в літню пору купають під душем. Корми для плідників заготовляють у місцевості, безпечній із заразних захворювань тварин. Перед початком роботи приміщення, у яких отримують, розріджують та розфасовують сперму, знезаражують бактерицидними лампами. Перед взяттям сперми плідників та підставних тварин чистять за допомогою щітки чи пилососу, особливо миють черевну стінку; калитку та препуцій обмивають теплим содовим розчином (1,5-2 %) і витирають насухо стерильною індивідуальною серветкою чи туалетним папером. При чищенні плідників особливу увагу звертають на нижню черевну стінку. Окремі спеціалісти радять промивати порожнину препуція антимікробними розчинами, проте при частому застосуванні цього прийому знижуються бактерицидні властивості слизової оболонки препуція. Тому промивання порожнини препуція 3 %-им розчином перекису водню, розчином фуразолідону 1: 10000 чи фурациліну 1: 5000 проводять один раз у декаду в спеціально відведеному місці з добрим стоком рідини. Після одержання сперми при необхідності видаляють з поверхні препуція вазелін 3 %-им розчином соди і витирають стерильною серветкою. Це важливо тому, що до залишків вазеліну на препуції прилипають різні механічні домішки (особливо, коли для підстилки використовують опилки), що можуть викликати баланопостит. За кожним плідником закріплюють по дві штучні вагіни і кожен еякулят беруть на окремо підготовану вагіну. Не дозволяється застосовувати одну і ту ж вагіну для одержання декількох еякулятів, оскільки при цьому сперма сильно забруднюється. Перед використанням штучні вагіни стерилізують у автоклаві, кип'ятять в дистильованій воді або, як виняток, знезаражують 96 %-им спиртом-ректифікатом. Перед стерилізацією кип'ятінням чи автоклавуванням на обидва кінці складеної вимитої вагіни одягають полотняні ковпаки або закривають обидва отвори приладу пергаментним папером, зафіксованим гумовими кільцями. Якщо після знезаражування внутрішня поверхня вагіни залишається вологою, її витирають насухо стерильною серветкою. Бугаям і баранам підвішують за передніми кінцівками перед препуцієм свіжови- митий, висушений і прогладжений фартух для попередження попадання пилу та мікроорганізмів з шерсті підставної тварини у сперму. Під час стрибка плідника фартух застилає круп підставної тварини і перешкоджає забрудненню статевого члена. Особливу увагу приділяють чистоті і стерильності спермоприймача. Одноразові поліетиленові спермоприймачі разом зі штучними вагінами стерилізують автоклаву- ванням при 105 °С протягом 30 хвилин. У Чехії для цього користуються спеціальним стерилізатором з ультрафіолетовим випромінювачем, розміщеним усередині трубчастого тримача, на який фіксують вагіну. Один раз у квартал проводять ветеринарно-санітарну оцінку сперми і змивів з пре- пуція шляхом дослідження їх на наявність мікробних тіл і колі-титр. До використання допускається сперма, що містить не більше 5 тисяч мікробних тіл в 1 мл при колі-ти- трі не більше 0,3 для сперми бугая, та 0,1 для сперми барана, кнура та жеребця. До і після кожного отримання сперми від кнура чучело миють теплою водою з милом і витирають насухо чистим рушником. Задню частину чучела протирають тампоном, змоченим у розчині фурациліну (1: 5000). Штучне осіменіння тварин в Україні, як і у інших країнах, проводять як спермою, отриманою від власних плідників, так і спермою, завезеною з-за кордону. Згідно наказів № 39 від 20.10.99 та № 52 від 27.09.02 Головного Державного інспектора ветеринарної медицини України дозволяється імпорт сперми бугаїв лише з підприємств, вільних від губкоподібної енцефалопатії великої рогатої худоби і скрепі овець, бруцельозу, лейкозу, туберкульозу, паратуберкульозу - протягом останніх 3-х років, інфекційного ринотрахеїту, вірусної діареї, трихомонозу, кампілобактеріозу, лептоспірозу, бесноітіозу, інфекційного кератокон'юнктивіту - протягом останніх 12-ти місяців. За 30 днів перед отриманням сперми бугаїв досліджують на туберкульоз, пара- туберкульозний ентерит, бруцельоз, лептоспіроз, лейкоз, блутанг, інфекційний ріно- трахеїт, вірусну діарею, трихомоноз, кампілобактеріоз, хламідіоз та інші інфекційні хвороби (залежно від епізоотичної ситуації). Не допускається наявності у спермі патогенних і токсикогенних мікроорганізмів та клітин крові. Бугаї-плідники, від яких одержують сперму для постачання на експорт, повинні бути не вакциновані проти бруцельозу. Сперма баранів-плідників повинна бути отриманою на підприємстві та адміністративній території, офіційно вільних від заразних хвороб тварин, у тому числі: губкопо- дібної енцефалопатії великої рогатої худоби і скрепі овець, меді-віспи, аденоматозу, блутангу, артриту-енцефаліту кіз - протягом останніх 3-х років, ку-лихоманки, африканської чуми свиней, ящуру, чуми дрібних жуйних, кампілобактеріозу, лістеріозу, інфекційної агалактії, інфекційного маститу - протягом 12 місяців на території країни. Барани-плідники не повинні бути вакциновані проти бруцельозу і епідидиміту. Сперма кнурів повинна надходити з підприємства і адміністративної території, офіційно вільних від заразних хвороб тварин, у тому числі: африканської чуми свиней - протягом останніх 3 років, ящуру, везикулярної хвороби свиней - протягом останніх 12 місяців; туберкульозу, бруцельозу, трихінельозу, репродуктивно-респіраторного синдрому свиней - протягом 3 років у господарстві. Кнури-плідники, від яких одержують сперму, не повинні бути вакциновані проти класичної чуми свиней. Сперма жеребців має надходити з підприємств та адміністративних територій, офіційно вільних від заразних хвороб тварин, у тому числі: інфекційних енцефало- міелітів коней усіх типів, вірусного артеріїту, африканської чуми коней - протягом останніх 3 років на території країни, ящуру, африканської чуми свиней, везикулярного стоматиту, грипу коней - протягом останніх 12 місяців на території країни; парувальної хвороби (трипаносома Авансі), піроплазмозу (бабезія Кабані) і таліозу (бабе- зія Екві) - протягом останніх 12 місяців на адміністративній території; інфекційного метриту коней - протягом останніх 12 місяців на території господарства; ринопнев- монії, інфекційної анемії і віспи коней - протягом останніх 3 місяців на території господарства. Жеребці-плідники, від яких беруть сперму, не повинні бути вакциновані проти ринопневмонії. Два рази на рік на племпідприємстві проводять диспансерне обстеження плідників у відповідності з планом протиепізоотичних заходів, проводять ветеринарно-санітарні заходи. При виникненні у плідників заразної хвороби, що може передаватися через осіменіння, запаси чи залишки отриманої від них сперми підлягають знищенню. Поводження з плідниками має бути спокійним, лагідним, але наполегливим, упевненим. Середовища, що застосовуються для розрідження сперми, повинні бути стерильними. Курячі яйця, жовтки яких використовуються для приготування середовищ, заготовляють у господарствах, безпечних по заразних хворобах. Розріджують і розфасовують сперму у стерильних умовах. Для попередження розвитку мікроорганізмів до середовищ додають сануючі препарати. Один раз на місяць на станції проводять ветеринарно-санітарний день. У цей день проводять клінічний огляд плідників, розчистку та обрізання у них копит, взяття проб сперми, слизу та змивів з препуція для перевірки їх мікробного забруднення та колі- титру, проводять дезінфекцію приміщень, інвентаря та предметів догляду. До роботи з штучного осіменіння тварин допускаються лише особи, що мають ветеринарну чи зоотехнічну освіту і пройшли відповідну підготовку. На пунктах штучного осіменіння необхідно суворо дотримуватися ветеринарно- санітарних правил. Всі приміщення пункту, обладнання та навколишню територію слід утримувати в чистоті та порядку, регулярно їх дезінфікувати непахучими дезза- собами. При вході у манеж має бути дезкилимок, який щоденно зволожують 20 %-им розчином їдкого натрію. Посуд, прилади та інструменти, що використовуються при осіменінні, мають бути чистими та стерильними. Оператор повинен працювати у чистому білому халаті, ковпаку чи косинці, після осіменіння кожної тварини повинен мити руки з милом і протирати їх тампоном, зволоженим 70°-им спиртом. Щоденно після закінчення роботи необхідно очистити і помити гарячим 2-3 %-им розчином соди станок, прибрати усі приміщення пункту і помити підлогу манежу. При приведенні на пункт хворої тварини чи виявленні її під час осіменіння необхідно припинити осіменіння, помити та продезінфікувати станок, та приміщення манежу і повідомити про це ветеринарну лікарню. Для осіменіння тварин у карантинованих та оголошених небезпечними господарствах користуються лише привізною спермою, яку доставляють у разових термосах, що не підлягають поверненню. Питання для самоконтролю 1. Як визначають оптимальний час для осіменіння корів та телиць? 2. Які методи застосовують для штучного осіменіння корів та телиць? 3. В чому суть цервікального осіменіння корів з ректальною фіксацією шийки матки? 4. Які особливості мано-цервікального осіменіння корів? 5. Як проводять візо-цервікальне осіменіння корів та телиць? 6. Як виявляють овець у стані охоти? 7. Якими методами користуються при штучному осіменінні овець? 8. Як визначають оптимальний час для осіменіння свиноматок? 9. Якими методами штучного осіменіння користуються у свинарстві? 10. Як визначають оптимальний час для осіменіння кобил? ЗАПЛІДНЕННЯ Запліднення - складний біологічний процес, що полягає у проникненні спермія у цитоплазму яйцеклітини, об'єднанні їх ядер з утворенням нового типу клітини - зиготи (гр. zygotos - з'єднані разом), з якої шляхом відповідної трансформації в геніталіях самки розвивається плід з властивими йому індивідуальними особливостями росту, розвитку та продуктивності. Пересування сперміїв у статевих органах самки. Запліднення відбувається у верхній третині яйцепроводу, у якому виділяють лійку, ампулу, перешийок (істмус) та інтрамуральний відділ. Перешийок є місцем природного резервування сперміїв, а ампула - місцем запліднення. Введені у статеві шляхи самки спермії для участі у заплідненні повинні пройти у ампулоподібне розширення яйцепроводів. Яким чином вони долають цю відстань? Ще з часів А. Левенгука, коли йому вперше вдалося побачити під мікроскопом рухливих сперміїв, у науці тривалий час існувала думка, що назустріч яйцеклітині спермії просуваються виключно за рахунок власного руху. Проте з часом ця думка змінилася. У процесі еволюції у ссавців виробилися пристосування, що сприяють зустрічі чоловічих та жіночих статевих клітин. Це, перш за все, відповідна морфологія та фізіологія статевої системи, морфологія та фізіологія гамет. Якщо, наприклад, співставити рух сперміїв тварин із зовнішнім і внутрішнім осіменінням, то у перших нормальним є коловий рух, оскільки при прямолінійному русі вони будуть розходитися у різні боки від місця знаходження яйцеклітини, тоді як у других саме прямолінійний поступальний рух буде сприяти переміщенню сперміїв від місця введення до місця запліднення. Проходженню сперміїв уздовж статевих шляхів самки сприяє їх здатність до реотаксису - руху проти течії. Це було доведено експериментально: нанесені на розтягнену поверхню розрізаного яйцепроводу спермії рухалися по слизовій оболонці, долаючи успішно опір війок та течії рідини. Осіменіння свійських тварин проводиться під час тічки, виділювані при цьому секрети статевих органів самки рухаються у протилежному напрямку і сприяють реотаксису сперміїв, як у піхві, так і у яйцепроводах. Проте одним реотаксисом не можна пояснити швидкого проникнення сперміїв у яйцепроводи. У тварин з матковим типом осіменіння цьому сприяють особливості статевого акту. Великий об'єм еякуляту, заповнюючи просвіт рогів матки, сприяє швидкому наближенню сперміїв до яйцепроводу. У тварин з піхвовим типом осіме- ніння з великої кількості введених сперміїв (4-5 млрд) у шийку матки проникає лише 0,05-0,5 % (4-20 млн). Велика складчастість слизової оболонки шийки матки та густий в'язкий секрет затримують тут сперміїв. Істотну роль у їх просуванні по статевих органах самки відіграє моторика матки та яйцепроводів, яка різко зростає з настанням тічки та охоти. Її підсилює виділюваний задньою долею гіпофіза окситоцин. Крім того, у спермі окремих видів тварин, зокрема у секреті простати, є простагландини, що володіють утеротонічною дією. Нарешті, після еякуляції у спермі утворюється ацетилхолін, що підсилює моторику матки. Сильні ритмічні скорочення поздовжніх м'язів матки супроводжуються розслабленням циркулярних м'язів, що приводить до скорочення матки і розкриття цервікального каналу. Позитивно позначається на моториці матки присутність самця. Як установив М. Полянцев, перистальтичні та антиперистальтичні скорочення матки у корів, найкраще виражені під час статевого збудження, підсилюються при наближенні бугая. Моторику матки також підсилює попадання у піхву корів секретів додаткових статевих залоз бугая. У дослідах на кобилах Г. В. Паршутін установив, що при підведенні жеребця до кобили у стані охоти її матка, що була до цього розслабленою, починає скорочуватися, спочатку скорочуються циркулярні м'язи, а через хвилину - поздовжні (циркулярні при цьому розслаблюються), внаслідок чого у порожнині матки наступає розрідження, що продовжується біля 3-6 хвилин. Завдяки такій засмоктуючій дії сперма досягає верхівки рогів матки протягом декількох секунд. При наступному розслабленні поздовжніх та скороченні циркулярних м'язів основна частина еякуляту засмоктується ще глибше. Добре виражена засмоктуюча дія матки у свиней. Скорочення матки у них стимулюється окситоцином, що виділяється рефлекторно з гіпофіза у відповідь на подразнення рецепторів геніталій під час статевого акту чи штучного осіменіння. Якщо під час осіменіння корови або зразу після нього її ударити чи крикнути на неї, то адреналін, який при цьому виділяється, гальмує дію окситоцину, знижує скорочення матки і затримує просування сперміїв по статевих шляхах. У 60-х роках минулого сторіччя група англійських біологів довела, що у жуйних у відповідь на подразнення рецепторів статевих органів під час нормально проведеного осіменіння внаслідок антиперистальтичних скорочень та засмоктуючої дії матки відбувається "механічне закидання сперміїв" з цервікального каналу до верхівок рогів матки. Для цього потрібно всього 3-5 хвилин. Отже, просування сперміїв по статевих шляхах самки відбувається за рахунок реотаксису, динаміки статевого акту та скорочень матки. У яйцепроводі додаються ще рухи війчастого епітелію та зростання тиску черевних стінок. З цього можна зробити висновок, що швидкість переміщення сперміїв по статевих шляхах самки залежить від виду осіменіння, рухливості сперміїв, реакції самки на присутність самця, подразнення рецепторів геніталій під час статевого акту чи штучного осіменіння. Швидкість переміщення сперміїв з місця введення до яйцепроводів у різних ссавців складає від 15 сек. до 1-3 годин. Є дані, що у корови вони досягають яйцепроводів через 2,5 хв. після осіменіння, тоді як В. Нагаєв виявляв їх лише через 3-14 годин. У овець одні дослідники виявляли їх через 0,5-1 годину, інші - через 6 хв. після осіменіння. У кобил спермії з'являються у яйцепроводі через 30-60 хв., а у свиней при природному осіменінні спермії досягали яйцепроводів через 15 хв., проникали у них через 0,5-1 годину, а при штучному осіменінні - досягали яйцепроводів через 45 хв., виявлялися у них через 1,5-2 години після осіменіння. У собак виявляли сперміїв у яйцепроводі через 25 сек. від початку еякуляції. Проте у яйцепроводи попадає лише частина сперміїв, більшість їх затримується у складках шийки матки. Частина сперміїв попадає у вивідні протоки маткових залоз, інші аглютинуються. Нарешті, вхід у яйцепровід розкривається лише під кінець охоти. На швидкість проходження сперміями до яйцепроводу впливає стан статевих шляхів та метод осіменіння. Яйцепровід, як і уся статева система самки, перебуває під нейрогуморальним контролем, парадоксальність якого полягає у тому, що у передовуляційну фазу спермії рухаються у напрямку ампули яйцепроводів, а після овуляції - у протилежному напрямку. Найкращим часом для досягнення ампул є період охоти. Спермії, що до- сягли перешийка яйцепроводу, скупчуються тут у передовуляційну фазу. Є дані, що вони можуть навіть впадати тут у стан анабіозу і перебувати в ньому до 18 годин. Після овуляції під впливом фолікулярної рідини їх рухливість поновлюється. Згідно Т. Бішопа і А. Грея, при температурі 37 °С спермій бугая здійснює 9 ударів за сек., просуваючись при кожному ударі на 8,3 мкм, а за одну хв. - на 4 482 мкм. Рух сперміїв по яйцепроводах забезпечується головним чином за рахунок рухів війчастого епітелію яйцепроводів та власних рухів сперміїв. Після овуляції погіршуються умови для просування сперміїв до верхівок яйцепроводів, цервікальний канал звужується, стає звивистим, скорочення матки стають перистальтичними. Згідно Р. Мартиненка, у свиней антиперистальтичні скорочення матки поступово наростають і досягають найбільшої інтенсивності через 24 години від початку охоти. Виживання гамет у різних ділянках статевих органів. Результативність осі- меніння буває тим вищою, чим менше часу пройде між уведенням сперми у статеві органи самки і овуляцією, яка у окремих тварин не співпадає з охотою, коли проводиться осіменіння. Більше того, у великої рогатої худоби овуляція відбувається через 15 годин після закінчення охоти. Ігнорування цими особливостями статевого циклу приводить до того, що нерідко осіменіння проводять передчасно чи запізно, спермії попадають у яйцепроводи або задовго до овуляції, або після неї, коли яйцеклітина або ще не може, або вже не може з'єднатися зі спермієм. Тому важливо знати, з одного боку, особливості виживання сперміїв у різних ділянках статевого апарату по періодах статевого циклу, а з іншого, оптимальний час осіменіння кожного виду тварин. Наявні дані наукових досліджень показують, що найгірші умови для виживання сперміїв у піхві, де вони зберігають рухливість 2-6 годин (у кобил 4-4,5, корів 1-6). Це зв'язане з низьким рН середовища, активним фагоцитозом, наявністю тут мікроорганізмів, домішок клітин крові. При виникненні запальних процесів у піхві виживання тут сперміїв значно знижується, наприклад, при вагініті у корів спермії гинуть протягом першої години, у кіз - через 1 годину 45 хв. після введення. Дані про виживання сперміїв у матці досить розбіжні. Одні вважають його досить високим, інші, навпаки, низьким, пояснюючи це наявністю тут, особливо у тварин з матковим типом осіменіння, лужного секрету, який активує рухливість сперміїв і вкорочує їх виживання. Хоча є дані, що виживання сперміїв у матці свиней та кобил, особливо у їх верхівках, сягає до 30-48 годин. У яйцепроводах виживання сперміїв знижується до 6-9 годин. Цікаві дані отримано у дослідах на кролях. Як відомо, овуляція у кролиць наступає приблизно через 10 годин після коїтусу. При штучному осіменіння кролиць високоякісною спермою з подальшим їх спаровуванням з вазектомованим самцем через 8 годин вагітність наступала у 90,9 % випадків. Якщо коїтус з вазектомованим самцем проводили через 20 годин після штучного осіменіння, то запліднення наступало лише у 12,9 % випадків. Нарешті, при коїтусі через 30 годин після осіменіння вагітність не наступала. Тобто, виживання сперміїв у статевих органах кролиці зберігається до 40 годин. Найвище виживання сперміїв у шийці матки, особливо у самок з піхвовим типом природного осіменіння. Тут спермії зберігають запліднюючу здатність найдовше, наприклад, у корови - до 30 годин, вівці - до 48 годин, хоча основна маса їх проходить через цервікальний канал досить швидко (15-50 хв). Тривалому зберіганню тут сперміїв сприяє велика складчастість слизової оболонки та наявність густого слизу із слабо кислою реакцією. Дослідження цервікального слизу показали, що він складається з глікопротеїду та плазми. Міцели глюкопротеїду розміщені паралельно і між ними можуть вільно просуватися спермії з прямолінійно-поступальним рухом. Такому розташуванню міцел глікопротеїду сприяє поява у слизі високих концентрацій естрогенів (і низьких - прогестерону) під час тічки та охоти. У лютеїнову фазу слиз стає в'язким, розщепленим, що значно перешкоджає руху сперміїв. Завдяки цьому, а також під впливом секретів додаткових статевих залоз під час охоти, активні спермії звільняються тут від мертвих та мікроорганізмів. Якщо ж осіменіння проводять після закінчення охоти, то спермії гинуть тут уже через 2-4 години, не досягаючи рогів матки. Отже, на всьому шляху від місця введення сперміїв до місця запліднення відбувається "відсів" слабших сперміїв (елімінація). Першим "фільтром" для сперміїв є цервікальний слиз. Далі відбувається відсів сперміїв з ферментними дефектами. Спермії, що проникли у матку, проштовхуються антиперистальтичними скороченнями її до верхівок яйцепроводів. Тривалість антиперистальтичних скорочень матки 6-8 годин. Яйцепровід розкривається під час овуляції і пропускає у корів та телиць 1 000 сперміїв, у овець - 500. У яйцепроводах рух сперміїв активізується фертилізином фолікулярної рідини, що забезпечує синхронність входження у зону запліднення яйця і спермія. Виживання сперміїв у статевих органах самки, збереження ними тут запліднюючої здатності визначаються фізіологічним станом самки, який у свою чергу піддається змінам у зв'язку з умовами утримання та годівлі тварин. Так, якщо осіменіння тварин проводиться не під час охоти, то вже через декілька хвилин спермії піддаються аглютинації і гинуть. В. Паршутін установив, що у статевих органах кобил з низькою вгодованістю спермії гинуть вже через 12-24 години, тоді як у кобил з середньою та доброю вгодованістю вони виживали до 48 годин. Окремі токсини, лікарські та інші речовини можуть проникати з кров'яного русла у статеві органи і виділювані ними секрети і позначатися на виживанні та запліднюючій здатності сперміїв. У дослідах з введенням лабораторним самкам per os свинцю, нікотину, фосфору відмічено значне скорочення виживання сперміїв, у 50 % випадків виникали аборти. Знижували також виживання сперміїв введення самкам миш'яку, ртуті, колоїдального срібла та заліза. Таким чином, виживання сперміїв у статевих органах самки визначається багатьма факторами, на першому місці серед яких слід поставити фізіологічний стан статевих органів та загальний стан здоров'я самки; зміни фізіологічного стану статевих органів, виникнення хворобливого стану чи, навпаки, одужання; неповноцінна чи, навпаки, повноцінна годівля викликають відповідні зміни у окремих ланках обміну речовин, характері і якості статевих секретів, тічкового слизу, що в свою чергу погіршує чи поліпшує виживання тут сперміїв. Безумовно, воно у свою чергу залежить від якості самої сперми, яка у прямій мірі залежить від фізіологічного стану плідника, стану його здоров'я, умов утримання, характеру годівлі та експлуатації. У птахів спермії скупчуються у складках та криптах лійки яйцепроводу і зберігають запліднюючу здатність протягом 2-3-х тижнів. У окремих рептилій та риб їх виявляли живими у спеціальних сперміотеках протягом декількох місяців і навіть років. Проте ефективність осіменіння залежить не лише від сперміїв, а й від виживання та біологічної активності яйцеклітини. А ці риси, на жаль, зберігаються у яйцеклітини значно коротше, ніж у сперміїв. Це, по-перше, зв'язане з тим, що яйцеклітина виходить з фолікула не повністю дозрілою, а у стані овоцита ІІ порядку. Другий поділ дозрівання яйцеклітини відбувається зразу після проникнення у неї спермія як наслідок активування яйця, що супроводжується двома морфологічними перетвореннями - відновленням мейозу та кортикальною реакцією. Якщо ж запліднення не наступило, то яйцеклітина швидко старіє і гине. По-друге, прозора оболонка яйцеклітини, на відміну від оболонки спермія, не проникна для поживних речовин, її енергетичні запаси досить обмежені. Згідно А. Лопиріна, старіння яйця супроводжується відповідними біохімічними змінами, які несприятливо позначаються на наступному онтогенезі. Тому при визначенні тривалості життя яйцеклітини слід враховувати не лише період, протягом якого можливе її запліднення (він складає для яйцеклітини близько 12-ти годин), але й час, протягом якого зберігається потенціальна здатність яйця до подальшого нормального онтогенезу, який, мабуть, не перевищує 5 годин. Під час овуляції яйцеклітина, разом з клітинами променевого епітелію (кумулюс) виходить у простір, оточений пристінковою очеревиною таза та широкою черевною зв'язкою, у якій відкривається своїм широким кінцем яйцепровід. У багатьох тварин кайма яйцепроводу тісно зв'язана з яєчником; внаслідок наповнення її судин кров'ю під час охоти вона стає щільною і притягується до яєчника, і навіть охоплює його. Згідно О. Квасницького, на місці розриву фолікула виявляється прозора драглиста маса, яка легко виймається з фолікула у вигляді тонкого ніжного тяжа, у середині якого знаходиться яйцеклітина, оточена променевим вінцем. Війчастими рухами клітин яйцепроводу цей тяж втягується у його просвіт. У тих же тварин, де кайма слабо розвинена (корова, вівця, коза), проникненню яйцеклітини у яйцепровід сприяють потоки рідини з черевної порожнини у матку. Як установив В. Груздєв, ці потоки скеровуються у напрямку матки коливальними рухами війчастого епітелію. Як тільки овоцит входить у яйцепровід, війковий епітелій, що раніше коливався у напрямку ампули яйцепроводу, тепер починає рухатися у зворотному напрямку, тобто від яєчника до матки. Переміщенню з потоками рідини яйцеклітини у яйцепровід сприяє також підвищення внутрішньочеревного тиску під час овуляції (К. Скробанський). Проте, це не виключає можливості деякої міграції яйцеклітини у черевній порожнині. У жінок, наприклад, у більшості випадків трубної вагітності овуляція відбувається у правому яєчнику, а плід розвивається у лівій трубі, що пояснюють зовнішньою (черевною) чи внутрішньою (матково-трубною) міграцією яйцеклітини. Отже, після овуляції яйцеклітина може деякий час плавати у капілярних щілинах між петлями кишечника, або ж переміщатися через тіло матки з одного яйцепроводу в інший. Істотний вплив на моторику яйцепроводів мають стероїдні гормони, катехоламі- ни, окситоцин та інші речовини. Яйцеклітина переміщується по ампулі яйцепроводу рухами епітелію, а у ділянці перешийка - також за рахунок скорочень його мускульного шару. До овуляції тут переважають антиперистальтичні, а після овуляції - перистальтичні скорочення. Керуючись даними з фізіології овуляції, у тому числі множинної, можна зробити висновок, що просування яйцеклітин по яйцепроводах триває від однієї до трьох діб, проте здатність до запліднення вони зберігають у межах 4-6 годин. Проникнення спермія у яйцеклітину та розвиток зиготи. Яйцеклітина, що вийшла з фолікула, кулястої форми, у ній розрізняють ядро та цитоплазму (ооплазму), оточені жовтковою оболонкою (оолемою) та прозорою оболонкою. Ззовні вона оточена клітинами променевого вінця, з'єднаними між собою в'язкими драглями, основу яких складає гіалуронова кислота (рис. 39). Для запліднення яйцеклітина повинна звільнитися від навколишніх клітин. У 1931 р. Ямане виявив у сперміях фермент гіалуронідазу, що розчиняє гіалуронові драглі. Вважають, що місія гіалуронідази зводиться до деполімеризації гіалуронової кислоти, руйнування міжклітинного ма- триксу, який з'єднує клітини променевого вінця, щоб дозволити окремому спермію пройти між фолікулярними клітинами до яйця. Згідно Остіна та Бішопа, для проникнення спермія у яйцеклітину не потрібно повного оголення її від клітин променевого вінця, а лише виникнення між ними щілини. Рис. 39. Яйцеклітина. Виходячи з цих міркувань, ---------------------------------------------------------------- І. Соколовська запропонувала вважати звільнення яйцеклітини від променевого вінця (денудація) першою стадією запліднення, хоча більшість учених вважають цей феномен підготовчою стадією. Сам процес запліднення включає такі стадії: 1) проникнення спермія крізь прозору оболонку; 2) активування яйця і блокада поліспермії; 3) утворення чоловічого та жіночого пронуклеусів: 4) заміна пронуклеусів хромосомними групами; 5) об'єднання двох хромосомних груп (сингамія). Механізм проникнення спермія через прозору оболонку повністю не з'ясований. Одні автори вважають, що у ньому беруть участь багато сперміїв, частина з яких навіть можуть занурюватися голівками у прозору оболонку, інші ж затримуються на її поверхні. Прихильники протилежної точки зору вважають, що як тільки спермій пройде між фолікулярними клітинами і доторкнеться голівкою прозорої оболонки чи може навіть зіллється з нею боковою поверхнею голівки, під впливом виділюваного ним ферменту зоналізину у прозорій оболонці виникає щілина, крізь яку він проникає у яйцеклітину. Прозора оболонка (zonapellucida) - це густе плетиво глікопротеїдів, у петлях якого є вуглеводні компоненти - кортикальні гранули ZP1, ZP2 і ZP3, з яких останній (ZP3) є специфічним рецептором для спермія. Прикріпившись до нього, спермій виділяє фермент акрозин (акросомна реакція), під впливом якого у прозорій оболонці виникає канал для проникнення спермія. Далі вступає у дію другий глікопротеїд (ZP2), що викликає затвердіння прозорої оболонки і захищає яйцеклітину від проникнення інших сперміїв. Наступає "блок поліспермії". Під час проникання спермія у глибину цитоплазми яйця завершується другий поділ мейозу та дозрівання яйцеклітини, виділяється друге напрямне тільце (рис. 40). Момент доторкання голівки спермія до яйця є початковою ланкою у ланцюгу послідовних змін, що розпочинаються з активування яйцеклітини: у неї зростає проник У деяких видів тварин (хом'як, сука, вівця) реакція прозорої оболонки наступає дуже швидко, тоді як у інших (наприклад, у щурів) вона сповільнена. У кролів не відмічено реакції прозорої оболонки, тому у пе- ривітеліновому просторі у них може скупчуватися багато сперміїв. Описані випадки проникнення у яйцеклітину кролиці до 200 "зайвих" сперміїв. Перивітеліновий простір відносно малий, тому голівка спермія, що проник сюди, торкається жовткової оболонки і наче прилипає до неї. Після деякої паузи спермій проникає крізь жовткову оболонку у цитоплазму. Вважають, що цей процес нага- --------------------------------------------------------------------------------------------------- дує фагоцитоз або захоплення амебою часток їжі. Він суворо специфічний, у ньому може брати участь лише спермій свого виду. Відбуваються зміни голівки спермія, вона з лопатоподібної стає циліндричною. Велику роль у цих змінах спермія і яйця відіграє секрет яйцепроводу, наявні у ньому інгібітори ферментів акрозину та трипсину, рівень яких зростає перед овуляцією, тоді значно знижується під час овуляції і знову зростає у наступні 1-2 дні. Ці інгібітори ферментів сприяють настанню у певний час овуляції і забезпечують захист ферментів сперміїв, що розпадаються. Спермії, що зосередилися у верхній третині яйцепроводу, перебувають у стані анабіозу і лише за декілька годин до овуляції піддаються капацитації
Зразу після проникнення спермія крізь жовткову оболонку його голівка відокремлюється від джгутика, збільшується в об'ємі (приблизно у 20 разів) і рухається у напрямку ядра яйцеклітини. Воно трансформується у жіночий пронуклеус, а ядро (голівка) спермія - у чоловічий пронуклеус. Чоловічі та жіночі пронуклеуси зближуються, зменшуються у об'ємі і втрачають свої оболонки, перетворюючись у хромосомні набори. Кінцевою стадією запліднення є сингамія - об'єднання хромосомних наборів самця і самки у одне ядро і виникнення нової клітини - зиготи з геномом нового індивіда. Центріолі, що були у шийці спермія, зразу утворюють веретено, у якому розташовуються хромосоми яйця і спермія, і зигота виявляється готовою до початку поділу. Дослідження Thibault показали, що запліднення - це не одномоментний процес. Тривалість перших двох стадій (проникнення спермія крізь прозору оболонку та зміни його голівки) становить в середньому по 3 години, третьої (перетворення голівки на чоловічий пронуклеус) - 4-5 годин, четвертої (зближення пронуклеусів) - 8-9, п'ятої (об'єднання пронуклеусів) - 10-12 годин. Таким чином, запліднення виконує подвійну роль: по-перше, воно виводить яйце із загальмованого стану, а, по-друге, воно приводить до утворення диплоїдної клітини зиготи з повним набором хромосом батьківської та материнської спадковості. Поділ зиготи торкається лише ядра при збереженні цілості прозорої оболонки. Внаслідок першого поділу зиготи утворений зародок має два бластомери. Кожне наступне покоління бластомерів виявляється меншим попереднього, тому цей поділ називають дробленням, що триває у корів та телиць близько 8-ми діб. Приблизно через кожних 24 години кількість бластомерів подвоюється, хоча у ссавців уже на ранніх стадіях ця чіткість може порушуватися. Новоутворені бластомери не розходяться на боки, а залишаються компакт
|
|||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 517; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.224.32.243 (0.014 с.) |