Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Локализация функций в коре большого мозгаСодержание книги
Похожие статьи вашей тематики
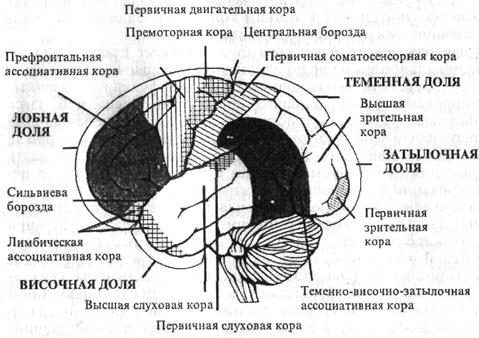
Поиск на нашем сайте А. Общая характеристика. В определенных участках коры большого мозга сосредоточены нейроны, воспринимающие определенный вид раздражителя: свет - затылочная область, звук - височная доля и т.д. Однако после удаления классических проекционных зон (слуховых, зрительных) условные рефлексы на соответствующие раздражители частично сохраняются. Согласно И.П.Павлову, в коре большого мозга имеются «ядро» анализатора (корковый конец) и «рассеянные» по всей коре нейроны. Современная концепция локализации функций базируется на принципе многофункциональности (но не равноценности) корковых полей. Свойство мульти-функциональности позволяет той или иной корковой структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную, генетически присущую ей функцию (О.С.Адрианов). Степень мультифункциональности различных корковых структур неодинакова. В полях ассоциативной коры она выше, чем в первичных сенсорных полях. В основе мультифункциональности лежат многоканальность поступления в кору мозга афферентного возбуждения, перекрытия афферентных возбуждений, особенно на таламическом и корковом уровнях; модулирующее влияние различных структур, например неспецифических ядер таламуса. ба-эальных ганглиев, на корковые функции; взаимодействие корково-подкорковых и межкорковых путей проведения возбуждения. С помощью микроэлектродной техники удалось зарегистрировать в различных областях коры большого мозга активность специфических нейронов, отвечающих на стимулы только определенной сенсорной модальности (только на свет, только на звук и т.п.), т.е. имеется множественное представительство функций в коре большого мозга. В детском возрасте наблюдается динамическая локализация функций, сущность которой заключается ъ том, что после удаления одной из зон коры по медицинским показаниям ее функция восстанавливается за счет других зон. Двигательные функции также распределены на значительных площадях коры большого мозга, нейроны, аксоны которых формируют пирамидный тракт и расположены не только в моторных областях, но и за их пределами. Наименее дифференцированными в функциональном отношении являются ассоциативные области, обладающие особо выраженными свойствами пластичности и взаимозамещаемости. В настоящее время принято подразделение коры на сенсорные, двигательные и ассоциативные (неспецифические) зоны. Б. Сенсорные зоны коры (проекционная кора, или корковые отделы анализаторов, по И.П.Павлову). В эти зоны поступает сенсорная информация. Они расположены преимущественно в теменной, височной и затылочной долях. Восходящие пути в сенсорную кору идут в основном от релейных сенсорных ядер таламуса. Поля 1, 2, 3 постцентральной области составляют основное корковое ядро кожного анализатора. Вместе с полем 1 поле 3 является первичной, а поле 2 - вторичной проекционной зоной кожного анализатора. Эфферентными волокнами постцентральная область связана с подкорковыми и стволовыми образованиями, с прецентральной и другими областями коры большого мозга. Первичные сенсорные зоны - это области сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма (ядра анализаторов, по И.П.Павлову). Они состоят главным образом из мономодальных нейронов и формируют ощущения одного качества. В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, их рецепторных полей. Вокруг первичных сенсорных зон находятся менее локализованные вторичные сенсорные зоны, нейроны которых отвечают на действие нескольких раздражителей, т.е. они полимодальны. Важнейшей сенсорной областью являются теменная кора постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий, которую обозначают как соматосенсорная область I. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и чувствительности опорно-двигательного аппарата - от мышечных, суставных, сухожильных рецепторов (рис. 5.13). Кроме соматосенсорной области I выделяют меньших размеров соматосенсорную область II, расположенную на границе пересечения центральной борозды с верхним краем височной доли, в глубине латеральной борозды. Степень локализации частей тела здесь выражена в меньшей степени. Хорошо изученной первичной проекционной зоной является слуховая кора (поля 41, 42, 52), которая расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). К проекционной коре височной доли относится также центр вестибулярного анализатора в верхней и средней височных извилинах (поля 20 и 21). В затылочной коре расположена первичная зрительная область (кора части клиновидной извилины и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки.
Рис. 5.13. Схема чувствительного и двигательного гомункулусов (по У.Пенфильду, Т.Расмуссену). Разрез полушарий во фронтальной плоскости, а - проекция общей чувствительности в коре постцентральной извилины; б- проекция двигательной системы в коре предцснтральной извилины Каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения. Около поля 17 расположена кора вторичной зрительной области (поля 18 и 19). Нейроны этих зонполимодальны и отвечают не только на световые, но и на тактильные и слуховые раздражители. В данной зрительной области происходит синтез различных видов чувствительности, возникают более сложные зрительные образы, и осуществляется их распознавание. Центр обонятельного анализатора находится в филогенетически наиболее древней части коры мозга - в крючке и аммоновом роге (поле Па, е), он обеспечивает проекционную функцию, а также хранение и распознавание обонятельных образов. Центр вкусового анализатора располагается в ближайшем соседстве с центром обонятельного анализатора, т.е. в крючке и аммоновом роге, но, кроме того, в самом нижнем отделе задней центральной извилины (поле 43), а также в островке. Как и обонятельный анализатор, центр обеспечивает проекционную функцию, хранение и распознавание вкусовых образов. Кзади от верхних отделов задней центральной извилины локализуется центр, обеспечивающий способность узнавания собственного тела, его частей, их пропорций и взаимоположения (поле 7). В нижних отделах передней и задней центральных извилин располагается центр анализатора интероцеппшвных импульсов внутренних органов и сосудов. Центр имеет тесные связи с подкорковыми вегетативными образованиями. В непосредственной близости от центра Вернике (средняя треть верхней височной извилины - поле 22) имеется центр, обеспечивающий распознавание музыкальных звуков, мелодий. На границе височной, затылочной и теменной долей располагается центр анализатора письменной речи (поле 39), который тесно связан с центром Вернике височной доли, с центром зрительного анализатора затылочной доли, а также с центрами теменной доли. Центр чтения обеспечивает распознавание и хранение образов письменной речи. Основная часть информации об окружающей среде и внутренней среде организма, поступившая в сенсорную кору, передается для дальнейшей ее обработки в ассоциативную кору, после чего инициируется (в случае необходимости) поведенческая реакция с обязательным участием двигательной коры. В. Двигательные зоны коры находятся в основном в прецен-тральной извилине (поля 4 и 6) и парацентральной дольке на медиальной поверхности полушария. Выделяют первичную и вторичную области - поля 4 и 6. Эти поля моторные, но по своей характеристике, согласно исследованиям Института мозга, они разные. В первичной моторной коре (поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (см. рис. 5.13). Основной закономерностью топографического представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков двигательной коры. Поле 4 полностью занято центрами изолированных движений, ноле 6 - лишь частично (подполе 6а), Сохранность поля 4 оказывается необходимой для получения движений при раздражении как поля 4, так и поля 6. У новорожденного поле 4 является практически зрелым. Раздражение первичной моторной коры вызывает сокращение мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральным). При поражении этой корковой зоны утрачивается способность к тонким координированным движениям конечностями и особенно пальцами рук. Вторичная двигательная кора (поле 6) имеет главенствующее функциональное значение по отношению к первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием и координацией произвольных движений. Здесь в наибольшей степени регистрируется медленно нарастающий отрицательный потенциал готовности, возникающий примерно за 1с до начала движения. Кора поля 6 получает основную часть импульсации от базальных ганглиев и мозжечка, участвует в перекодировании информации о сложных движениях. Раздражение коры поля 6 вызывает сложные координированные движения, например поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения сгибателей или разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле 44), обеспечивающие речевой праксис, а также музыкальный моторный центр (поле 45), обеспечивающий тональность речи, способность петь. Нижняя часть поля 6 (подполе бор), расположенная в области покрышки, реагирует на электроток ритмическими жевательными движениями. Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, от базальных ганглиев и мозжечка. Основным эфферентным выходом двигательной коры на стволовые и спинальные моторные центры являются пирамидные клетки V слоя. Основные доли коры большого мозга представлены на рис. 5.14. Другие корковые центры. Центр поворота глаз и головы в противоположную сторону расположен в средней лобной извилине впремоторной области (поля 8, 9). Работа этого центра тесно связана с системой заднего продольного пучка, вестибулярными ядрами, образованиями стриопаллидарной системы, участвующей в регуляции торсии, а также с корковым отделом зрительного анализатора (поле 17). В задних отделах верхней лобной извилины представлен центр, дающий начало лобно-мостомозжечковому пути (поле 8). Эта область коры больших полушарий участвует в обеспечении координации движений, связанных с прямохождением, сохранением равновесия стоя, сидя и регулирует работу противоположного полушария мозжечка.
средней лобной извилины, в непосредственной близости от проекционной корковой зоны руки (поле 6). Центр обеспечивает автоматизм письма и функционально связан с центром Брока. Распределение парциальных центров движений в преценграль-ной области и центров общей чувствительности в постцентральной области таково, что соматотопическая проекция в коре может быть представлена как единая для моторной и сенсорной функций. Имеется полное совпадение моторных центров, соответствующих определенным территориям иннервации, с аналогичными сенсорными центрами, распределенными внутри постцентральной области. Установление соматотопической проекции в коре полушария мозга человека имеет огромное значение для правильного диагносцирования локализации патологического процесса. У человека при частичной экстирпации двигательной коры, как и при патологическом процессе, немедленно возникает вялый паралич соответствующей конечности. Корковые дефекты при патологических процессах редко бывают настолько обширны, чтобы привести к длительному выпадению функций всей моторной зоны. Ассоциативные области коры Ассоциативные области коры (неспецифическая, межсенсорная, межанализаторная кора) включают участки новой коры большого мозга, которые расположены вокруг проекционных зон и рядом с двигательными зонами, но не выполняют непосредственно чувствительных или двигательных функций; им нельзя приписывать преимущественно сенсорных или двигательных функций, нейроны этих зон обладают большими способностями к обучению. Границы указанных областей обозначены недостаточно четко. Ассоциативная кора является филогенетически наиболее молодой частью новой коры, получившей наибольшее развитие у приматов и человека. У человека она составляет около 50% всей коры, или 70% неокортекса. Термин «ассоциативная кора» возник в связи с существовавшим представлением о том, что эти зоны за счет проходящих через них кортико-кортикальных соединений связывают двигательные зоны и одновременно служат субстратом высших психических функций. К ассоциативной коре в узком смысле, слова относят те-менно-височно-затылочную префронтальную кору лобных долей и лимбические ассоциативные зоны. Нейроны ассоциативной коры являются полисенсорными (полимодальными): они отвечают, как правило, не на один (как нейроны первичных зон), а на несколько раздражителей: один и тот же нейрон может возбуждаться при раздражении слуховых, зрительных, кожных и других рецепторов. Полисенсорность нейронов ассоциативной коры создается кортико-кортикальнымисвязями с разными проекционными зонами, связями с ассоциативными ядрами таламуса. В результате этого ассоциативная кора представляет собой своеобразный коллектор различных сенсорных возбуждений и участвует в интеграции сенсорной информации и в обеспечении взаимодействия сенсорных и моторных областей коры. В настоящее время с учетом таламокортикальных проекций выделяют две основные ассоциативные системы мозга: таламотеменную и таламолобную. Таламотеменная система представлена ассоциативными зонами теменной коры, получающими основные афферентные входы от задней группы ассоциативных ядер таламуса. Теменная ассоциативная кора имеет эфферентные выходы на ядра таламуса и гипоталамуса, в моторную кору и ядра экстрапирамидной системы. Основными функциями таламотеменной системы являются гнозис и праксис. Под гнозисом понимают функцию различных видов узнавания - формы, величины, значения предметов, понимание речи, познание процессов, закономерностей и др. К гностическим функциям относится оценка пространственных отношений (например, взаимного расположения предметов). В теменной коре выделяют центр стереогнозиса, обеспечивающий способность узнавания предметов на ощупь. Вариантом гностической функции является формирование в сознании трехмерной модели тела («схемы тела»). Под праксисом понимают целенаправленное действие. Центр праксиса находится в надкраевой извилине левого полушария, он обеспечивает хранение и реализацию программы двигательных автоматизированных актов. Таламолобная система представлена ассоциативными зонами лобной коры, имеющими основной афферентный вход от ассоциативного медиодорсального ядра таламуса, других подкорковых ядер. Основная роль лобной ассоциативной коры сводится к инициации базовых системных механизмов формирования функциональных систем целенаправленных поведенческих актов (П.К.Анохин). Префронтальная область играет главную роль в выработке стратегии поведения. Нарушение этой функции особенно заметно, когда необходимо быстро изменить действие и когда между постановкой задачи и началом ее решения проходит некоторое время, т.е. успевают накопиться раздражители, требующие правильного включения в целостную поведенческую реакцию. Некоторые ассоциативные центры (например, стереогнозиса, праксиса) включают в себя и участки височной коры. В височной коре расположен слуховой центр речи Вернике, находящийся в задних отделах верхней височной извилины левого полушария. Этот центр обеспечивает речевой гнозис - распознание и хране- ние устной речи, как собственной, так и чужой. В средней части верхней височной извилины находится центр распознания музыкальных звуков и их сочетаний. На границе височной, теменной и затылочной долей находится центр чтения, обеспечивающий распознание и хранение образов. Высказывается мнение о целесообразности выделить и третью — таламовисочную - систему. НЕЙРОФИЗИОЛОГИЧЕСКИЕ
|
||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 855; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.3 (0.015 с.) |