Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Симпатическая нервная системаСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте А. Иннервируемые органы. В отличие от парасимпатических нервов, которые выходят из различных отделов ЦНС, все симпатические нервы выходят из спинного мозга (нейроны расположены в боковых рогах, сегменты Суш - Ьн - ядро Якобсона, или ядро парамедианное дорсальное) и иннервнруют все органы и ткани организма (рис. 1.2). Симпатические нервы выходят в составе передних корешков и, отделившись от них, идут в виде белых соединительных ветвей к симпатическому стволу. Это преганглионарные мякотные (мислиновые) волокна. Часть из них прерывается в узлах симпатического ствола, другая часть проходит, не прерываясь. От симпатического ствола отходят два нервных пути. Первый путь - серые соединительные ветви, представляющие собой постганглионарные симпатические волокна. Они вступают в спинномозговые (соматические) нервы, иннервирует лимфатические и кровеносные сосуды, железы, мышцы, поднимающие волосы, мышцы туловища и конечностей, скелетную мускулатуру, все органы и ткани. Второй путь - нервы, идущие непосредственно к внутренним органам и сосудам. Небольшая часть волокон этих нервов является постганглионарными, большая - преганглионарными. Последние образуют синапсы на клетках превертеб-ральных, а также шейных, нижних поясничных, крестцовых и копчиковых ганглиев. От нейронов этих ганглиев отходят постганглионарные волокна, иннервирующие внутренние органы и ткани организма. Б. Локализация симпатических центров. В последнем шейном и двух первых грудных сегментах спинного мозга расположен спино-цилиарный центр, осуществляющий регуляцию функции гладких мышц глаза, в том числе мышцы, расширяющей зрачок. Эффекторные нейроны этого пути лежат в краниальном шейном симпатическом ганглии. От Н-1У грудных сегментов спинного мозга отходят симпатические волокна, иннервирующие слюнные железы. В первых пяти грудных сегментах спинного мозга расположены симпатические нейроны, иннервирующие сердце и бронхи. Ганглионар-ные клетки этого пути лежат в основном в звездчатом ганглии или, реже, в узлах пограничного симпатического ствола. Сосуды и потовые железы иннервируются всеми симпатическими преганглионарными нейронами спинного мозга. Таким образом, симпатические нервы регулируют функции всех органов и тканей организма, включая ЦНС и сенсорные рецепторы. Норадренергические нейроны голубого пятна по своим морфологическим, биохимическим и электрофизиологическим свойствам весьма сходны с периферическими нервными клетками. Есть данные о том, что многие норадренергические волокна, выходящие из нижних частей ствола мозга, иннервируют артериолы и капилляры коры полушарий большого мозга. Подобные нейроны могут участвовать в регуляции кровотока через кору, их можно рассматривать как центральный отдел симпатической нервной системы.
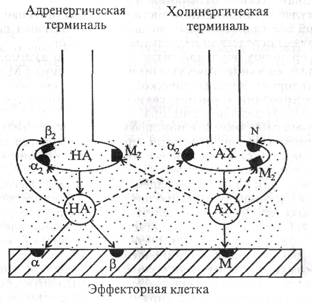
ганглиев можно зарегистрировать тормозные - гиперполяризационные -потенциалы. Вызываются они вставочными адренергическими клетками, регулирующими возбудимость ганглионарных нейронов. 2. В постганглионарных симпатических окончаниях главным медиатором является норадрепалин - его около 90% (адреналина около 7%, дофамина около 3%). Синтезируется норадреналин из аминокислоты тирозина в аксоне ганглионарного нейрона, в основном в терминальных симпатических окончаниях, примерно 1% - в теле нейрона. Поскольку в окончаниях симпатических ганглионарных нейронов выделяется норадреналин, эти нейроны называют адренергическими (рис. 1.3).
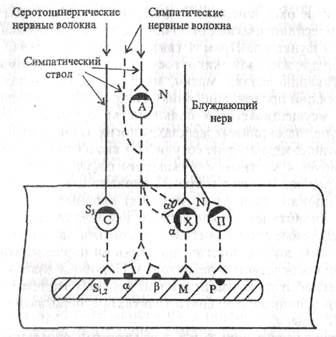
деляется из симпатических терминален, варикоз. Мобильный норадреналин пополняется из стабильного фонда. Наиболее значим в процессах выделения медиатора везикулярный экзоцитоз, состоящий из нескольких этапов. Первый этап - сопряжение деполяризации и секреции - реализируется с помощью Са2+, который проникает в пресинаптические нервные окончания через медленные и быстрые кальцевые каналы. На место выделившихся везикул медиатора транспортируются по филаментам с затратой энергии новые гранулы медиатора. Источник энергии - АТФ. Если Са2+ обеспечивает попадание медиатора в экзоцитозный карман, то происходит выдавливание медиатора с помощью актомиозина. Выведение Са2+ из нервного окончания после его возбуждения осуществляется кальциевым насосом. Инактивация медиатора. Примерно 75-80% выделившегося в синаптическую щель норадреналина захватывается обратно пресинаптической мембраной и поступает в пузырьки. Не попавший в пузырьки норадреналин разрушается моноаминоксидазой (МАО), локализованной на пресинаптической мембране. Часть выделившегося в синаптическую щель и не связавшегося с пост- и пресинаптическими рецепторами норадреналина разрушается в области рецепторов постсинаптической мембраны ферментом катехол-О-метилтрансферазой (КОМТ) клеток-эффекторов, часть медиатора диффундирует в интерстиций и кровь. Вклад МАО и КОМТ в процессы инактивации выделившихся в синаптическую щель катехоламинов невелик, а доля участия этих ферментов в разных органах и тканях различна: в ЦНС действует в основном МАО, в периферической симпатической нервной системе - КОМТ. 3. Эффекторные рецепторы. Выделившийся из симпатических окончаний иорадрсналин действует на а- и (3-адренорецепторы, являющиеся гликопротеидами (см. рис. 1.3). Деление рецепторов основано на чувствительности их к различным фармакологическим препаратам: а-адренорецепторы блокируются фентолами-ном, а р-адренорецепторы - пропранололом. Оба типа рецепторов делятся на два подтипа: a1- и a2-, b1- и b2-адренорецепторы. Антагонистами a1-адренорецепторов являются празозин, дропе-ридол, антагонистами а2-адренорецепторов - раувольсин, йохимбин, b1-дренорецепторов - практолол, атенолол, b2-адренорецепторов - бутоксамин. Распределение a- и b-адренорецепторов в различных органах. В большинстве органов, реагирующих на катехоламины, содержатся a- и b -адренорецепторы, причем одна гладкая мышечная клетка может иметь оба рецептора (табл. 1.1). В чистом виде выделены из мембраны a1-адренорецепторы. Они представляют собой белок с молекулярной массой 9600 и обусловливают сокращение сосудов, матки, семявыносящего протока, семенных пузырьков, мышц, поднимающих волосы, и расслабление продольного и циркулярного мышечных слоев кишечника. Г. Связь симпатических окончаний с адренорецепторами. Считают, что симпатическую иннервацию имеют преимущественно a1 и b1-адренорецепторы (постсинаптические рецепторы). Активируются они в основном медиатором норадреналином a2 и b2-адренорецепторы, как правило, не имеют иннервации или она выражена слабо, они расположены вне синапсов на клетках-эффекторах и активируются циркулирующим в крови адреналином и диффундирующим норадреналином после его выделения пресинаптическими окончаниями. Эти адренорецепторы обнаружены также на пресинаптических окончаниях, где они выполняют регулирующую функцию. Прямая связь посредством синапса имеет место в сердце, жировой ткани, сосудах и в ряде гладкомышечных органов (мышцы зрачка, матки, мышца, поднимающая волосы, семявыносящий проток, кишечник). Непрямой тип регуляции, как известно, осуществляется в большинстве органов: в скелетной мускулатуре, эндокринных железах, многих экзокринных железах. В этих органах медиатор из сосудистых симпатических сплетений диффундирует к местным окружающим сосуд клеткам либо разносится кровью. Имеется и третий вариант связи симпатических (постганглионарных адренергических) нервных волокон - они образуют синаптические контакты с нейронами интраорганной нервной системы: либо с преганглионарными парасимпатическими волокнами, либо с постганглионарными парасимпатическими волокнами посредством а-рецепторов. Подобные взаимодействия симпатической и парасимпатической нервных систем установлены в органах пищеварительного тракта, предполагаются в сердце и мочевом пузыре. Д. Эффекты активации a- и b -адренорецепторов и механизмы их реализации. В физиологических условиях реакция какого-либо органа на адреналин и норадреналин, поступающих с кровью либо выделяющихся при возбуждении симпатических нервов, зависит от преобладания a- или b -адренорецепторов в данном органе и различной их возбудимости. Активация a1- и a2-адренорецепторов мембран клеток-эффекторов ведет к деполяризации их и повышению функции органа, в том числе сфинктеров пищеварительного тракта. Мышцы желудка и кишечника при этом расслабляются. Однако если симпатический ствол раздражается в грудной полости, то, как правило, регистрируются не тормозные, а стимуляторные реакции желудка и двенадцатиперстной кишки. Стимуляторные реакции осуществляется, по-видимому, с помощью серотонинерги-ческих нервных волокон, имеющихся в составе симпатических стволов (рис. 1.4). Активация b-адренорецепторов вызывает разнонаправленные эффекты в различных органах. Активация р-адренорецспторов в сердце (их назвали b1-рецепторами, они хорошо иннервированны) ведет к деполяризации и возбуждению кардиомиоцитов, увеличению частоты и силы сердечных сокращений, повышению автома-тии, облегчению атриовентрикулярной проводимости. При активации b-адренорецепторов эффекторных клеток гладкомышечных органов (их назвали b2 -адренорецепторами, они менее иннервированны) возникает пшерполяризация, наблюдается снижение функции органа, в частности расширение сосудов.
котехоламинов являются также цГМФ, диацилглицерин (ДАГ). При взаимодействии катехоламинов с рецепторами особо важную роль играет регуляторный G-белок. Этот белок играет важную роль в функционировании и других рецепторов: более 80 типов рецепторов связаны с ионными каналами или ферментами посредством G-белка. Е. Мозговой слой надпочечников. Этот слой представляет собой видоизмененный симпатический ганглий. Его клетки, с онтогенетической точки зрения, гомологичны ганглионарным симпатическим нейронам. Они содержат включения, окрашивающиеся в желто-коричневый цвет двухромовокислым калием, что и послужило поводом назвать их хромаффинными клетками. В виде скоплений хромаффинные клетки встречаются на поверхности аорты, в области каротидного синуса, среди клеток симпатических ганглиев. Преганглионарные волокна образуют на этих клетках, как и на хромаффинными клетках надпочечников, возбуждающие холинергиче-ские синапсы. Выделение катехоламинов из мозгового вещества надпочечников регулируется исключительно нервными влияниями (перерезка преганглионарных симпатических волокон прекращает секрецию катехоламинов). При возбуждении преганглионарных волокон у человека в кровоток обычно выбрасывается смесь катехоламинов, состоящая из адреналина (80-90%) и норадреналина (10-20%). Точки приложения для продуцируемых надпочечниками катехоламинов те же, что и у симпатической нервной системы, однако их действие более выражено, чем симпатических нервов, в областях с редкой адренергической иннервацией (в циркулярных и продольных мышцах кишечника, крупных артериях, матке). Взаимодействие катехоламинов с адренорецепторами вызывает различные эффекты у разных органов, в частности торможение функции пищеварительного тракта, улучшение процесса передачи в нервно-мышечных синапсах и увеличение силы сокращений скелетных мышц, увеличение частоты и силы сокращений сердца, расширение бронхов. Все это имеет важное приспособительное значение, обеспечивая мобилизацию систем организма при физическом и эмоциональном напряжениях. Катехоламины усиливают высвобождение свободных жирных кислот из подкожной жировой ткани и образование глюкозы и лактата из гликогена, необходимых клеткам организма при напряжении. Ускоренное расщепление углеводов осуществляется с помощью активации аденилатциклазы, стимулирующей образование цАМФ, который в свою очередь активирует фосфорилазу, расщепляющую гликоген - источник энергии. Таким образом, катехоламины мозгового слоя надпочечников можно рассматривать как метаболические гормоны. Не случайно симпатэктомированные животные не в состоянии осуществлять физические усилия, плохо переносят охлаждение и перегревание, с большим трудом справляются с кровотечением. У симпатэктомированных животных не бывает проявления характерных защитных реакций и показателей агрессивности: тахикардии, повышения артериального давления, расширения зрачков. Введение в организм ганглиозидов стимулирует рост новых синапсов, ускоряет реиннервацию после повреждения нервных волокон. Клетки, подобные хромаффинным мозгового слоя надпочечников, называют трансдуктороми, их короткие аксоны не имеют синаптических контактов с другими клетками, они выделяют в кровь свои биологически активные вещества, называемые также гормонами. К клеткам-трансдукторам относят, кромехромаффинных клеток, нейроны супраоптического и паравентри-кулярного ядер гипоталамуса, выделяющие вазопрессин и окситоцин; нейроны гипоталамуса, выделяющие в сосудистую систему факторы, регулирующие функцию гипофиза; клетки юкстагломерулярного аппарата почки, которые под влиянием посттангаионар-ных симпатических волокон выделяют в кровь ренин. Таким образом, симпатическая нервная система вместе с мозговым слоем надпочечников (симпатико-адреналовая система) активирует деятельность всего организма, мобилизует его защитные силы, обеспечивает выход крови из кровяных депо, поступление в кровь глюкозы, ферментов, усиливает метаболизм тканей, увеличивает расход энергии. Возбуждение симпатической нервной системы является пусковым звеном стрессорных эмоционально окрашенных реакций.
ждающий нерв) отходят к дыхательным органам, большей части пищеварительного тракта (до нисходящей ободочной кишки), сердцу, печени, поджелудочной железе, почке. Парасимпатические нервы сакрального отдела спинного мозга иннервируют нисходящую часть ободочной кишки и тазовые органы (прямая кишка, мочевой пузырь, половые органы). Парасимпатической иннервации не имеют скелетные мышцы, матка, мозг, значительное большинство кровеносных сосудов (кожи, органов брюшной полости, скелетных мышц), органы чувств и мозговое вещество надпочечников. Парасимпатические ганглии и отдельные нейроны расположены внутри органов, а в тазовой области и в области головы - в непосредственной близости от органов. От нервных клеток парасимпатических ганглиев идут короткие постганглионарные парасимпатические волокна, иннервирующие все перечисленные ранее органы; преганглионарные волокна обычно длинные (у симпатической нервной системы наоборот: преганглионарные - короткие, постганглионарные — длинные). Б. Медиаторы и рецепторы. 1. Проведение возбуждения с преганглионарного парасимпатического волокна па эффекторный нейрон осуществляется с помощью ацетилхолина (см. рис. 1.3). Медиатор действует на М-холинорецептор постсинаптической мембраны ганглионарного нейрона. 2. Постганглионарное волокно свое влияние на эффекторную клетку передает также с помощью ацетилхолина, который в парасимпатических терминалиях находится в трех фондах (пулах): 1) стабильный, прочно связанный с белком ацетилхолин, не готовый к освобождению; 2) мобилизационный, менее прочно связанный и пригодный к освобождению ацетилхолин; 3) готовый к освобождению ацетилхолин (активный медиатор), который освобождается квантами спонтанно и при поступлении к нервному окончанию потенциалов действия. Активный медиатор локализуется в прилежащих к мембране пузырьках. Пузырьки с медиатором находятся в движении, и по мере расходования активного медиатора к пресинаптической мембране поступают новые пузырьки с активным ацетилхолином. Освобождению квантов медиатора способствует ион Са2+. Ацетилхолин синтезируется в цитоплазме окончаний холинергических нейронов, депонируется в везикулах по нескольку тысяч в каждой. Инактивация медиатора. Выделившийся в синаптическую щель ацетилхолин, как и в любом другом синапсе, не весь используется для передачи сигнала. В отличие от симпатической нервной системы основная часть ацетилхолина разрушается ферментом ацетил-холинэстеразой с образованием холина и уксусной кислоты, которые захватываются пресинаптической мембраной и вновь используются для синтеза ацетилхолина. Значительно меньшая часть медиатора диффундирует в интерстиций и кровь. Обратного захвата нерасщепленного ацетилхолина нервными окончаниями не происходит. По поводу локализации ацетилхолинэстеразы существуют две точки зрения: согласно одной из них, фермент фиксирован на постсинаптической мембране вблизи холинорецепторов, согласно другой - на базальной мембране. Последняя представляет собой тонкую сеть коллагена и гликозоаминогликинов между пре- и постсинаптическими мембранами. 3. Эффекторные рецепторы. На клетки-эффекторы ацетилхолин действует с помощью М-холинорецепторов (см. рис. 1.3), которые свое название получили от мускарина - токсина мухомора, который возбуждает эти рецепторы и вызывает такой же эффект, как и ацетилхолин. Мускаринолодобный эффект ацетилхолина был открыт позже. М-холинорецепторы имеются также на симпатических и парасимпатических окончаниях, в коре головного мозга, ретикулярной формации. По чувствительности к различным фармакологическим препаратам выделяют М1—М4-холинорецеп-торы. М1-холинорецепторы локализуются в вегетативных ганглиях и ЦНС, М2-холинорецепторы - в сердце, на пресинаптических окончаниях, гладких мышцах пищеварительного тракта. На гладких мышцах расположены также Мз-холинорецепторы, имеются они и в большинстве экзокринных желез. М4-холиноре-цепторы изучены мало. Некоторые симпатические нервные волокна (иннервирующие потовые железы, симпатические волокна, вызывающие расширение сосудов скелетных мышц) также являются холинергическими. Убедительного объяснения этому до настоящего времени нет. Сам по себе факт передачи сигнала с постганглионарных симпатических волокон посредством ацетилхолина установлен давно, многократно подтвержден в эксперименте и не вызывает сомнений. Однако постганглионарное симпатическое волокно является аксоном адренергического нейрона, в его окончаниях синтезируется норадреналин. Следовательно, либо симпатические преганглионарные во-локна на своем пути где-то образуют синапсы с холинергическими нейронами, либо постганглионарные симпатические волокна образуют синапсы на поштанглионарных парасимпатических термина-лях и, возбуждая их, обеспечивают выделение ацетилхолина. В. Эффекты возбуждения парасимпатической нервной системы (активации эффекторных М-холинорецепторов) проявляются сокращением гладких мыщц пищеварительного тракта - усиливается перистальтика, сокращением мыщц бронхов - их просвет суживается, мочевого пузыря, сфинктера зрачка - зрачок суживается, сокращением ресничной мышцы глаза - хрусталик становится более выпуклым. Одновременно тормозится деятельность сердца, расширяются сосуды половых и некоторых других органов. Активация парасимпатических нервов сопровождается увеличением секреции всех желез, иннервируемых ими: пищеварительного тракта, слюнных, трахеи и бронхов. Сфинктеры желудочно-кишечного тракта и мочевого пузыря расслабляются. Парасимпатическая нервная система способствует восстановлению гомеостазиса, обеспечивая трофотропный эффект. Примером стимулирующего влияния парасимпатических нервов на гладкую мускулатуру является усиление сокращений желудка и кишечника при раздражении блуждающего нерва. Однако более слабое раздражение этого же нерва нередко вызывает противоположный эффект - угнетение сокращений органа. Механизмы этого феномена до настоящего времени не раскрыты. Эффекты экзогенного ацетилхолина те же, что и парасимпатической нервной системы. Г. Механизм действия ацетилхолина. Стимулирующее влияние ацетилхолина на любой орган осуществляется, во-первых, посредством изменения характера электрических процессов (вызов возбуждения его клеток); во-вторых, посредством биохимических реакций с помощью вторых посредников: инозитолтрифосфата, Са2+. Тормозной эффект ацанияхолина также возникает в результате изменения характера электрических процессов (гиперполярйзация клеток эффектора). При этом активируются вторые посредники: гуанилатцик-лаза - циклический гуанозинмонофосфат (ГЦ - цГМФ). Основные эффекты стимуляции парасимпатических нервов приведены в табл. 1.1, схема эфферентного отдела вегетативной нервной системы-на рис. 1.4. Регуляция функции синапсов Эффективность передачи в синапсе вегетативной нервной системы (ВИС) зависит от количества медиатора, выделяемого в синаптическую щель, которое определяется числом импульсов, пришедших к терминали: с увеличением импульсации выделение медиатора увеличивается до определенного значения, при ослаблении импульсации - уменьшается. Кроме того, важную роль в регуляции выделения медиатора в синаптическую щель лостганглионарными окончаниями ВНС играют пресинаптические адрено- и холиноре-цепторы, имеющиеся на самих окончаниях. Эти рецепторы реализуют свое влияние по принципу отрицательной обратной связи, вызывающей противоположные эффекты: малые порции медиатора стимулируют его выделение, большие - угнетают. А. Регуляция выделения норадреналина. На симпатических постганглионарных окончаниях имеются а2- и b2-адреноре-цепторы и М2-холинорецепторы. Активация М2-холинорецепторов и a2-адренорецепторов тормозит, b2-адренорецепторов -стимулирует освобождение медиатора (рис. 1.5).
мозят освобождение норадреналина из пре-синаптических окончаний. Активация пресинаптических М2-холинорецепторов осуществляется ацетилхолином, выделяющимся из рядом расположенных парасимпатических окончаний. На окончаниях преганглионарных симпатических волокон имеются рецепторы к серотонину, гистамину, кортикостероидам и К-холи-норецепторы, активация которых облегчает синаптическую передачу N-холинорецепторы, рецепторы дофамина, напротив, обеспечивают ингибирование секреции ацетилхолина преганглионарными симпатическими окончаниями. Б. Регуляция освобождения ацетилхолина из пресинаптичсских окончаний постганглионарных волокон. Эта регуляция осуществляется также посредством нескольких механизмов. 1. По принципу обратной связи - с помощью взаимодействия выделившегося в синаптическую щель ацетилхолина с М2- и N-холинорецепторами пресинаптической мембраны: связывание с М2-холинорецепторами тормозит, связывание с N-холинорецептора-ми усиливает выделение медиатора. 2. Ретроингибирование с помощью АТФ обеспечивается пури-норецепторами пресинаптической терминали. Большое количество АТФ выделяется в синаптическую щель под влиянием ацетилхолина из эффекторной клетки. 3. Тормозящее влияние норадреналина, выделяющегося из рядом расположенного симпатического синапса, обеспечивается в парасимпатическом синапсе действием на a2-рецепторы пресинаптической мембраны, что угнетает освобождение ацетилхолина, - перекрестное взаимное торможение активности симпатических и парасимпатических терминалей. В. Эффективность передачи в синапсе. Передача в синапсе регулируется также числом рецепторов на гюстсинаптической мембране: при уменьшении выделения медиатора синтез рецепторов постсинаптической мембраной возрастает, чувствительность ее повышается (сенситизация); в случае увеличения выхода медиатора синтез рецепторов уменьшается, чувствительность постсинаптической мембраны снижается (десенситизация). Одно и то же биологически активное вещество может выполнять функции медиатора и модулятора. Так, например, ацетилхолин и норадреналин, выделяемые из постганглионарных нервных окон- чаний, действуют не только на рецепторы эффекторных клеток, но одновременно регулируют выделение медиатора самими нервными окончаниями - проявляется их модулирующая роль (см. раздел 1.1Л,Б). Некоторые вещества, по-видимому, играют роль только модуляторов функции нервных окончаний и эффекторных клеток. Наиболее важными нейромодуляторами являются кортикостерои-ды, половые гормоны, АКТГ, соматостатин, ангиотензин, эндор-фин, энкефалины, простагландины. Взаимодействие нейромодуля-торов с клеточными внесинаптическими рецепторами вызывает изменение либо проницаемости клеточной мембраны, либо клеточного метаболизма в результате их проникновения в цитоплазму клеток. В обоих случаях это ведет к усилению влияния нервных волокон на эффекторную клетку.
|
||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 1188; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.02 с.) |