Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механизм тщательного рефлексаСодержание книги
Похожие статьи вашей тематики
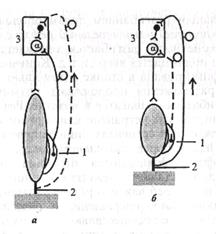
Поиск на нашем сайте А. Рефлекс возникает при однократном раздражении кожи конечности. Он выражается в сгибании этой конечности с одновременным дополнительным разгибанием противоположной задней конечности - перекрестный разгибателъпъш рефлекс (см. рис. 5.2, Б). Затем согнутая конечность разгибается, опускается вниз, разогнутая - сгибается и поднимается вверх и т.д. Конечности не касаются опоры (собака фиксирована в станке с помощью лямок) и без дополнительного раздражения продолжают ритмично поочередно сгибаться и разгибаться - «шагать» в воздухе. Рефлекс может продолжаться часами, так как устранено влияние коры большого мозга. При неповрежденных сегментах, иннервирующих передние конечности (Су-ТЬп), и более длительном раздражении задней конечности в рефлекс вовлекаются передние конечности. Это наблюдение свидетельствует о межсегментарной связи на уровне спинного мозга, играющей важную роль в ритмических движениях конечностей. Поочередное сокращение и расслабление мышц - сгибателей и разгибателей осуществляются в результате взаимодействия процессов возбуждения и торможения в соответствующих центрах спинного мозга под влиянием импульсов, поступающих в мозг от проприорецепторов. Б. Особая роль проприорецепторов в осуществлении шагательно-го рефлекса определяется их расположением. Мышечные веретена (мышечные рецепторы) расположены параллельно скелетной мышце — своими концами они крепятся к соединительнотканной оболочке (перимизий) пучка экстрафузальных (п!5и& - веретено) мышечных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5-1,0 мм (рис. 5.6). Поэтому, когда мышца расслабляется (удлиняется), растягиваются и мышечные рецепторы, что и ведет к их возбуждению (см. рис. 5.6, а). Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон, окруженных соединительнотканной капсулой. Длина волокон равна 4-7 мм, толщина составляет 15-30 мкм. Длина экстрафузальных мышечных волокон - от нескольких миллиметров до многих сантиметров, толщина составляет 50-100 мкм. Вокруг средней части мышечного веретена несколько раз обвивается окончание одного афферентного волокна. Многие мышечные веретена иннервируются и другим - более тонким - афферентным волокном, которое обвивает в виде спирали периферические участки мышечного рецептора. Импульсы от мышечных рецепторов возбуждают нейроны своего центра (гомонимные нейроны) и тормозят нейроны центра-антагониста. Ближе к концам мышечного веретена подходят двигательные нервные окончания, являющиеся аксонами у-мотонейронов спинного мозга. Их импульсация вызывает сокращение интрафузальных мышечных волокон (мышечных волокон рецептора), что ведет к возбуждению рецепторов.
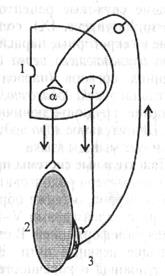
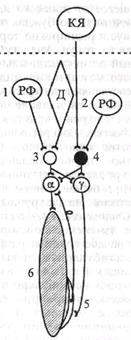
представляют собой безмиелиновые окончания толстого миелино-вого афферентного волокна (подойдя к капсуле рецепторов Гольджи, это волокно теряет миелиновую оболочку и делится на несколько окончаний). Сухожильные рецепторы крепятся к скелетной мышце последовательно, что обеспечивает их раздражение при натяжении сухожилия (см. рис, 5.6, б). Поэтому сухожильные рецепторы посылают в мозг информацию о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы - о том, что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц-сгибателей это возбуждение выражено слабее). В. Элемент шагательного рефлекса - поочередное сокращение и расслабление скелетной мышцы под влиянием импульсов от про-приорецепторов. поступающих в свой центр. Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от которых поступают к своим омотонейронам спинного мозга и возбуждают их (рис. 5.6, а). Далее сс-мотонейроны посылают импульсы к этой же скелетной мышце, вызывая ее сокращение. Как только мышца сократилась (рис. 5.6, b), возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), но начинают возбуждаться сухожильные рецепторы. Импульсы от рецепторов также поступают в первую очередь в свой центр спинного мозга, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение а-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление (удлинение) снова ведет к возбуждению мышечных веретен и а-мотонейронов - мышца вновь сокращается. Вследствие ее сокращения возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что вновь ведет к расслаблению скелетной мышцы и т.д. Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разгибателю. При этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу.
возбуждении центра мышцы-сгибателя тормозится центр мышцы-разгибателя. Если на одной ноге сокращены сгибатели (рис. 5.7, А), на другой ноге должны сокращаться разгибатели (рис. 5.7, Б), что обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров сгибателей и разгибателей. Регуляция тонуса мышц Тонус исследуют после поперечной перерезки спинного мозга. Сразу после перерезки в эксперименте или после травматического повреждения его у человека наблюдаются мышечная атония и отсутствие рефлексов (спинальный шок). Главная причина спинального шока - выключение влияния на спинной мозг вышележащих отделов ЦНС (повторная перерезка спинного мозга ниже первой после исчезновения шока повторно шок не вызывает). Спинальный шок у лягушек длится несколько минут, у собак - несколько дней, у человека - около 2 мес. После исчезновения спинального шока тонус мышц, иннервируемых посредством сегментов спинного мозга, которые находятся ниже перерезки (повреждения), резко повышается. Сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются. Механизм повышения тонуса мышц нижних конечностей в случае повреждения спинного мозга (после окончания спинального шока) отражает рис. 5.8. Гипертонус имеет рефлекторную природу - он развивается вследствие афферентной импульсации от мышечных рецепторов. Деафферентация, например, нижней конечности у лягушки ведет к исчезновению тонуса у нее (опыт Бронжеста). Афферентную импульсацшо от мышечных рецепторов по задним корешкам спинного мозга, которая ведет к возбуждению а-мо-тонейронов спинного мозга и сокращению скелетных мышц, обеспечивают три фактора: • растяжение и возбуждение мышечных рецепторов под влиянием сил гравитации конечности; • спонтанная активность мышечных рецепторов; • спонтанная активность у-мотонейронов, которая реализуется следующим образом. Возбуждение у-мотонейронов вызывает возбуждение и сокращение иннервируемых ими ин-трафузальных мышечных волокон, в результате чего увеличи- вается натяжение мышечного рецептора, поскольку концы его зафиксированы на скелетной мышце. Вследствие натяжения мышечного веретена раздражаются и возбуждаются его афферентные окончания(проприорецепто-ры), импульсы от которых поступают к а-мотонейро-нам, возбуждая их. В свою очередь а-мотонейроны посылают импульсы к скелетной мышце и вызывают ее постоянное (тоническое) сокращение. Мы рассмотрели рефлекторную и проводниковую функции изолированного спинного мозга. В нормальных условиях деятельность спинного мозга контролируется вышележащими отделами ЦНС посредством импульсации ко всем его нервным элементам.
VII) ызывает сокращение мимической мускулатуры. К ядрам преддверно-улиткового нерва (n. vestibulocochlearis, VIII) относятся: вестибулярные ядра, которые получают импульсы от вестибулорецепторов и участвуют в регуляции позы и равновесия тела; улитковые ядра, иннервирующие слуховые рецепторы. Ядра языкоглоточного нерва (n. glossopharyngeus, IX), содержащего двигательные, чувствительные и секреторные парасимпатические волокна. Ядра блуждающего нерва (n. vagus, X) регулируют функции внутренних органов (парасимпатический нерв). Вожбуждение двигательного ядра добавочного нерва (п. accessories, XI) вызывает сокращение грудино-ключично-сосцевидной и трапециевидной мышц. Двигательное ядро подъязычного нерва (n. hypoglossus, XII) иннервирует мышцы языка. Б. Двигательные системы продолговатого мозга и мостя. Из перечисленных ядер продолговатый мозг включает ядра 1Х-ХII пар черепных нервов, а также образования ретикулярной формации. Мост представлен ядрами V-VIII пар черепных нервов, а также ретикулярной формацией. В стволе мозга проходят восходящие и нисходящие нервные пути. Важную роль в регуляции тонуса мышц туловища и конечностей играют вестибулярные ядра, основным из которых является ядро Дейтерса. 1. Рефлексы, дуги которых замыкаются в продолговатом мозге и мосту, можно объединить в три основные группы: • жизненно важные вегетативные: дыхания, сердечно-сосудистой системы, пищеварительной системы, глотания; • защитные рефлексы: чиханья, кашля, мигания, слезоотделения, рвотный; • соматические рефлексы, участвующие в регуляции тонуса и двигательной активности мышц туловища, конечностей, шеи, лица. Рефлексы любого отдела ЦНС изучают после отделения вышележащих отделов мозга. У бульварного животного, у которого произведена перерезка ствола мозга между мостом и средним мозгом, сохраняются все рефлексы, в возникновении которых играют роль спинной мозг, а также указанные черепные нервы и их ядра. Особо важное значение в регуляции тонуса мышц туловища и конечностей имеет ядро Дейтерса. С его помощью осуществляется, в частности, лабиринтный рефлекс позы - повышение тонуса мышц-разгибателей при положении животного вверх ногами. Рефлекс в натуральных условиях направлен на сохранение позы. Он возникает с рецепторов вестибулярного аппарата (при его разрушении рефлекс исчезает). Проприорецепторы мышц шеи при изучении этого рефлекса должны быть выключены, что достигается путем гипсования шеи (исключаются сгибание шеи и раздражение проприорецепторов мышц шеи). 2. Общее состояние бульварного животного характеризуется тем, что произвольные движения с помощью мышц, иннервируемых отделами ЦНС, расположенными ниже перерезки, невозможны, тонус мышц-разгибателей сильно повышен, поэтому конечности напряженно выпрямлены, спина вогнута. Если такое животное поставить на ноги, оно будет стоять, как «скамейка». Такое состояние называют децеребрационной ригидностью; оно является следствием дисбаланса возбуждающих и тормозных влияний ядер ствола мозга на а- и у-мотонейроны спинного мозга, а также импульсации от мышечных рецепторов (рис. 5.9).
1. Характеристика рефлексов, осуществляемых средним мозгом (мезенцефальное животное). Они более совершенны, их набор значительно больше, тонус мышц близок к нормальному, поскольку сохраняется тормозящее влияние красного ядра на а- и у-мотонейроны спинного мозга, наблюдается незначительная ригидность (см. рис. 5.9). Руброспиналънъш тракт (тракт Монакова), которыйначинается от красного ядра покрышки среднего мозга и идет в спинной мозг, возбуждая а- и у - мотонейроны сгибателей, повышает их тонус и реципрокно тормозит мотонейроны мышц - разгибателей, снижая их тонус. Аналогично на мышечный тонус действуют латеральный ретикулоспинальный (от ретикулярной формации продолговатого мозга) и пирамидный кортикоспинальный тракты. Вместе они составляют латеральную нисходящую сгибательную систему. Таким образом, в стволе мозга имеются четыре главных парных моторных центров и трактов, регулирующих тонус мышц туловища и конечностей, - два ретикулоспинальных пути, руброспинальный путь и волокна ядра Дейтерса (см. рис, 5.9). Рефлексы мезенцефального животного изучил голландский физиолог Р.Магнус (1924). Соматические рефлексы, связанные с изменением положения тела (точнее - головы) и перемещением его в пространстве, Р. Магнус разделил соответственно на две группы - статические и статокинетические. 2. Статические рефлексы (перераспределение мышечного тонуса при изменении положения головы в покое) в свою очередь делятся на две группы: рефлексы позы (шейные и лабиринтные вестибулярные) и выпрямительные (выпрямление головы и туловища). Рефлекс выпрямления головы возникаете рецепторов вестибулярного аппарата, кожи и сетчатки. При этом возбуждаются проприорецепторы мышц шеи, и начинается выпрямление туловища. В осуществлении выпрямительных рефлексов принимают участие также и рецепторы кожи — дублирование рефлексогенных зон. При выключении рецепторов кожи или вестибулярного аппарата выпрямительные рефлексы не нарушаются. При выключении обеих названных рефлексогенных зон выпрямительные рефлексы не возникают. Для осуществления выпрямительных рефлексов, т.е. восстановления нарушенной позы, необходимо обязательное участие красных ядер, повреждение которых, как показано в эксперименте на животных, предотвращает возникновение выпрямительных рефлексов. Однако у человека и обезьян выпрямительные рефлексы осуществляются при обязательном участии коры большого мозга; при ее выключении выпрямительные рефлексы не возникают. При изменении положения головы появляются также рефлексы компенсаторного положения глаз, обеспечивающие правильное видение при различных положениях головы относительно тела и в пространстве. Например, если повернуть голову вправо, глазное яблоко поворачивается влево, как бы стремясь удержать первоначальное поле зрения. 3. Статокинетические рефлексы возникают при ускорениях прямолинейного и вращательного движений организма. Перераспределение тонуса направлено на преодоление действующих на человека ускорений и сохранение нормальной позы. Для их осуществления необходимо обязательное участие структур среднего мозга. Примером рефлекса прямолинейного ускорения является лифтный рефлекс. В начале быстрого подъема лифта (или при остановке движущегося вниз лифта) в нижних конечностях повышается тонус мышц-разгибателей. В начале опускания лифта (или при остановке движущегося вверх лифта) повышается тонус мышц-сгибателей. О рефлекторном характере изменения тонуса мышц конечностей при лифтном рефлексе свидетельствует его отсутствие у животных при выключении вестибулярных рецепторов разрушением лабиринта. Статокинетические рефлексы вращения включают мускулатуру тела и глазные мышцы. Движение глазных яблок (глазной нистагм) при этом способствует сохранению зрительной ориентации. Нистагм имеет две фазы. При ускорении вращения сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению. После этого они быстро отклоняются обратно (по ходу вращения). Медленное отклонение вызывается с рецепторов полукружных каналов, быстрый компонент связан с влиянием корковых центров (при наркозе он исчезает). При замедлении вращения, напротив, медленное движение глаз направлено в сторону вращения, а быстрое - против направления вращения. Движение глаз при нистагме (вправо-влево, вверх-вниз, круговые движения) зависит от возбуждения рецепторов соответственно горизонтального, сагиттального и фронтального полукружных каналов. Наряду с регуляцией мышечного тонуса, позы и равновесия структуры ствола головного мозга участвуют в контроле спинального автоматизма шагания и, следовательно, в контроле локомоции (например, ходьбы). Важное значение в осуществлении этой функции придают клинообразному ядру в задненижнем отделе среднего мозга, который обозначают как локомоторную область среднего мозга, имеющую эфферентный выход к ядрам моста. Произвольная локомоция без участия корковых нейронов невозможна. В условиях эксперимента при сохранении среднего мозга у животного возникает сторожевой рефлексов - повороты головы, глаз, ушей, туловища в сторону звука или света (настора-живание), при этом повышается тонус мышц-сгибателей (животное как бы готовится к прыжку). В случае действия светового раздражителя наблюдаются зрачковый рефлекс (сужение зрачка), аккомодация, дивергенция и конвергенция зрительных осей, чтоосуществляется с помощью передних холмиков четверохолмия (первичный зрительный центр) и III и IV пар черепных нервов. Задние холмики четверохолмия - первичные слуховые центры. 4. Функции голубого пятна. Оно расположено в среднем мозге, оказывает моделирующее влияние на двигательные механизмы спинного мозга. Аксоны норадренергических нейронов голубого пятна идут в переднем и боковом канатиках к а-мотонейронам передних рогов спинного мозга. Полагают, что норадреналин является тормозным медиатором для н-мотонейронов. Импульсная активность нейронов голубого пятна повышена в фазе быстрого сна, что выключает мышечный тонус и фазические движения мускулатуры туловища, конечностей. При одностороннем разрушении голубого пятна возникают вращательные движения в сторону, противоположную разрушению, что, как предполагают, связано с нарушением функциональных связей голубого пятна и базальных ганглиев. В электрофизиологических экспериментах было показано, что норадренергические нейроны голубого пятна оказывают ингибирующее действие почти на все иннервируемые ими образования ЦНС. Поскольку эти нейроны возбуждаются во время различных стрессовых воздействий, считают, что их ингибирующий эффект играет двоякую роль: 1) под их влиянием при стрессе нервная система предохраняется от перевозбуждения: 2) возбудимость клеток нервной системы поддерживается на постоянном уровне с целью оптимальной передачи сигналов. 5. Функции черной субстанции. Она участвует в регуляции мышечного тонуса, координации жевания и глотания, мелких движений конечностей (например, печатание на машинке, письмо), в осуществлении эмоций. В случае повреждения черной субстанции наблюдается бедность движений. Во всех двигательных реакциях и регуляции мышечного тонуса весьма важную роль играет ретикулярная формация, основная масса нейронов которой находится в стволе мозга.
|
||||||||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 568; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.016 с.) |