Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Глава вторая движения, параличи и парезыСодержание книги
Поиск на нашем сайте
Двигательная функция человека представляется чрезвычайно сложной. В осуществлении движений участвует целый ряд отделов нервной системы. В одних случаях движения примитивны, происходят непроизвольно по типу простого рефлекторного акта и осуществляются за счет деятельности сегментарного аппарата (спинного мозга, мозгового ствола). Примером таких простых автоматических движений у человека являются разобранные выше рефлексы. Существуют, однако, и более сложные автоматизмы, осуществляемые не только сегментарным аппаратом, но и более высокими отделами нервной системы с участием экстрапирамидных иннервации. Ряд еще более сложных движений осуществляется не только за счет иннервации экстрапирамидной системы и сегментарного аппарата, но и при известном участии коры головного мозга, например ходьба в обычных условиях. Наконец, существует многочисленная категория движений, при которых участие коры головного мозга является основным, а иннервации экстрапирамидные и сегментарные — лишь подсобными. Сюда относится ряд целенаправленных действий, осуществляющих сложные акты двигательного поведения человека(праксии). Движения, возникающие в результате корковых иннервации, выработанные в жизненном опыте и являющиеся, по существу, условными рефлексами, обычно называются «произвольными»; автоматические рефлекторные движения — «непроизвольными». Для осуществления «произвольного» движения необходимо, в частности, чтобы импульсы, возникающие в коре головного мозга, были проведены к мышце. Проведение импульса из коры происходит по цепи, состоящей из двух невронов: 1) центрального двигательного и 2) периферического двигательного неврона. Весь путь называется кортико-му-скулярным (tractus corticomuscularis), "^ Центральный двигательный неврон начинается от области коры, расположенной кпереди от роландовой борозды в передней центральной извилине, в задних отделах верхней и средней лобных извилин и в lob-ulus paracentralis. Нервные клетки, дающие волокна для иннервации отдельных мышечных групп, имеют расположение, обратное расположению частей человеческого тела: проекция движений нижней конечности — в верхних отделах передней центральной извилины, верхней конечности — в среднем ее
отделе, а головы, лица, языка, глотки и гортани - в нижнем. В заднем отделе верхней лобной извилины представлена проекция движений туловища и в заднем отделе средней лобной извилины - поворота головы и глаз в противоположную сторону. Иннервация мускулатуры перекрестная, т.е. правое полушарие связано с мускулатурой левой половины тела, а левое — с правой (перекрест проводников, о чем будет сказано ниже). Центральный двигательный неврон берет начало от больших пирамидных клеток Беца, расположенных в пятом слое коры проекционной двигательной зоны. Аксоны этих клеток идут от коры в глубь полушарий, проходят в составе волокон coronae radiatae белое вещество полушарий и двумя проводниками, т.е. tractus corticobulbaris или nuclearis (PNA) (от нижнего отдела передней центральной извилины) и tractus corticospinalis (от верхних двух третей извилины), проходят через внутреннюю капсулу (capsula interna). Внутренняя капсула расположена между большими ганглиями основания; она представляет собой полосу белого вещества, в котором расположены проводники, идущие от коры головного мозга к нижележащим отделам центральной нервной системы, и восходящие проводники, поднимающиеся к коре. Cepsula interna делится на переднее бедро (между nucleus caudatus и п. lentiformis), заднее бедро (между nucleus lentiformis и thalamus opticus) и колено (место перегиба, genu capsulae internae, см. рис. 85). Tractus corticonuclearis (центральные невроны для мускулатуры лица, языка, гортани и глотки) проходит в области колена внутренней капсулы; tractus corticospinalis, иначе называемый пирамидным пучком, расположен в передних двух третях заднего бедра внутренней капсулы (см. рис. 85, У). В направлении своем книзу оба названных проводника переходят из внутренней капсулы в ножки мозга, занимая средние две трети основания их. Tractus corticonuclearis расположен здесь медиально, tractus corticospinalis — латерально. В мосту (pons Varolii) двигательные проводники располагаются также в основании. Проходящими здесь поперечно и перекрещивающимися волокнами средних ножек мозжечка (tractus pontocerebellares) пирамидные пути разделяются на ряд отдельных пучков, сливающихся снова в общий проводник в продолговатом мозге. В продолговатом мозге пирамидные пучки находятся в основании, обособляясь в два валика, заметные на поверхности мозга, - пирамиды (см. рис. 76).
На границе продолговатого и спинного мозга пирамидные пучки (tractus corticospinalis) подвергаются неполному перекресту (decussatio pyramidum). Большая, перекрещенная часть пути переходит в боковой столб спинного мозга и называется основным, или латеральным, пирамидным пучком, меньшая, неперекрещенная часть проходит в передний столб спинного мозга и носит наименование прямого неперекрещенного пучка (см. рис. 27). Волокна tractus corticonuclearis заканчиваются в двигательных ядрах черепно-мозговых нервов, tractus corticospinalis — в передних рогах спинного мозга. Здесь импульсы с центральных двигательных невро-нов передаются на периферические. Периферические двигательные невроны для мускулатуры жевательной, лицевой, языка, гортани и глотки состоят из моторных клеток двигательных ядер черепных нервов с их аксонами, составляющими двигательные волокна корешков и нервов V, VII, IX, X и XII (тройничного, лицевого, языкоглоточного, блуждающего, подъязычного). Клетки периферических двигательных невронов для мускулатуры шеи, туловища, конечностей и промежности расположены в передних рогах спинного мозга; аксоны их выходят в составе передних двигательных корешков и составляют двигательные волокна периферических спинномозговых нервов. В настоящее время различают три вида периферических двигательных невронов: альфа большие-, альфа малые- и гамма-мотоневроны. Аксоны аль-фа-мотоневронов заканчиваются в мионевральных пластинках мышц, а тонкие аксоны гамма-невронов (рис. 9) идут к мышечным проприоцепторам (к интрафузальным волокнам мышечных веретен). Импульсы гамма-невронов повышают чувствительность проприоцепторов к давлению и растяжению мышц. Кора головного мозга, а также подкорковый и стволовый отделы мозга влияют на работу мышц как через альфа-мотоневроны, так и через гамма-петлю (гамма-неврон - передний корешок - мышечные проприоцепторы - зад-
Рис. 9. Альфа- и гамма-мотоневроны. / - гамма-мотоневрон; 2 — альфа-мотоневрон: 3 — мышечное веретено (рецептор); 4 — поперечнополосатая мышца. ний корешок — альфа-мотоневрон — мышечные волокна). Гамма-мотоневроны оказывают постоянное влияние на тонус поперечнополосатых мышц. Корковая иннервация мускулатуры - преимущественно перекрестная, так как волокна центральных двигательных невронов в большинстве своем переходят на противоположную сторону и устанавливают связи с противоположными ядрами черепных нервов и передними рогами спинного мозга. Перекрест волокон tractus corticospinalis происходит сразу на границе продолговатого и спинного мозга; волокна же tractus corticonuclearis подвергаются перекресту последовательно, по мере подхода их к соответствующим ядрам черепных нервов («надъядерный» перекрест). При поражении центральных двигательных путей расстройства движений, понятно, возникают в мускулатуре противоположной стороны тела, однако не во всей: ряд мышечных групп остается непораженным. Это объясняется наличием двусторонней корковой иннервации, существующей для глазодвигательных, жевательных мышц, мышц глотки, гортани, шеи, туловища и промежности. Суть в том, что к части двигательных ядер черепных нервов и к некоторым уровням передних рогов спинного мозга волокна центральных двигательных невронов подходят не только с противоположной стороны, но и со своей, обеспечивая, таким образом, подход импульсов из коры не только противоположного, но и своего полушария. Понятно, что при одностороннем центральном поражении выпадают функции только мышечных групп, односторонне (только из противоположного полушария) иннервируемых. Таковыми оказываются конечности, язык и нижний отдел лицевой мускулатуры, что мы и наблюдаем при так называемой гемиплегии; мускулатура же шеи, туловища, промежности и большая часть мышц, иннервируемых черепными нервами, остается при этом непораженной.
Для осуществления движения необходимо, чтобы двигательный импульс беспрепятственно был проведен из коры головного мозга к мышце. При перерыве кортико-мускулярного пути проведение импульса невозможно и соответствующая мускулатура оказывается в состоянии паралича. Неполная утрата движений (уменьшение их силы и объема) называется не параличом, а парезом. По своей распространенности параличи делятся на моноплегии (парализована одна конечность), гемиплегии (паралич одной половины тела), параплегии (поражение двух симметричных конечностей, верхних или нижних); тетраплегии (парализованы все четыре конечности). Параличи или парезы, вызванные поражением отдельных нервных стволов, обозначаются как паралич соответствующего нерва, например лучевого, локтевого и т.д. Аналогично этому различают параличи сплетений (плечевого, поясничного) или отдельных стволов их. При поражении центральных двигательных невронов возникает центральный паралич; при поражении периферических невронов — периферический. Общим для них является лишь самый факт паралича, в остальной симптоматология их резко различается. ПЕРИФЕРИЧЕСКИЙ ПАРАЛИЧ Периферический паралич является, как только что было сказано, результатом поражения периферических двигательных невронов, т.е! клеток передних рогов спинного мозга (или двигательных ядер черепных нервов), передних корешков и двигательных волокон спинномозговых и черепных периферических нервов (сведения о сегментарном аппарате, нервах и мышцах, участвующих в осуществлении некоторых движений, представлены в таблице 2). Этот тип параличей характеризуется утратой рефлексов, гипотонией и дегенеративной атрофией мышц, сопровождающейся так называемой реакцией перерождения. Утрата рефлексов (или ослабление их при неполном поражении) становится понятной, если мы вспомним, что периферический двигательный невроз является в то же время и центробежной, эфферентной частью рефлекторной дуги. При перерыве любого отдела последней рефлекторный акт невозможен или (при неполном перерыве) ослаблен. Атония или гипотония мышц объясняется также перерывом рефлекторной дуги, в результате чего мышца утрачивает свойственный ей постоянный, так называемый контрактильный, тонус, поддерживаемый в норме той же рефлекторной дугой с участием гамма-петли. Кроме того, атония может быть усилена возникающей атрофией мышечной массы. Атонические мышцы на ощупь дряблы, вялы, пассивные движения избыточны, суставы «разболтаны». Такое состояние мускулатуры дает основание называть периферический паралич также вялым, или атоническим.
Атрофия мышц возникает в результате разобщения с клеткой переднего рога, откуда по двигательному нервному волокну к мышце притекают нервнотрофические импульсы, стимулирующие нормальный обмен мышечной ткани. Наличие мышечных атрофии обусловливает еще одно определение периферического паралича - как атрофического. Атрофия мышц наступает вслед за перерождением и гибелью нервных двигательных волокон, происходит «денервация» мышцы. В итоге в нервах исчезают книзу от места перерыва двигательные волокна; в мышце развивается дегенеративный процесс, характеризующийся изменениями мышечных волокон, гибелью их, развитием жировой и соединительной ткани. Возникают характерные типичные для периферического паралича изменения электрических реакций пораженных нервов и мышц, называемые реакцией перерождения или дегенерации В норме при раздражении нерва гальваническим (при замыкании и размыкании) и фарадическим токами происходит сокращение ин-нервируемых им мышц; при раздражении теми же токами непосредственно самой мышцы также происходит ее сокращение причем на Таблица 2
Продолжение таблицы 2
Продолжение таблицы 2
Продолжение таблицы 2
гальванический ток оно возникает чрезвычайно быстро («молниеносно») и отличается тем, что катодозамыкательное сокращение больше, чем анодозамыкательное (КЗС > АЗС)1. При реакции перерождения (дегенерации) нерв не проводит тока к мышце, ибо двигательные центробежные волокна его перерождены и погибли; сама мышца денервирована и утрачивает способность сокращения на раздражение фарадическим током, сохраняя возбудимость только на гальванический. Но и это сокращение становится медленным («червеобразным»), причем большим может стать уже анодозамыкательное сокращение (АЗС > КЗС). Такое состояние называется полной реакцией перерождения, и наступает она на 12-15-й день после перерыва нерва или гибели клетки переднего рога. Бесспорным признаком денервации является вялый характер сокращения, а не изменения соотношения полюсов. Если сокращение остается живым, то даже при извращении формулы реакции перерождения нет (В.К.Рот). 1 Иными словами, для вызова сокращения мышцы на катоде требуется меньшая сила тока, чем на аноде. Таблица 3
При неполном поражении периферического двигательного неврона может наступить частичная реакция перерождения, когда возбудимость нерва на оба тока не утрачена, а лишь ослаблена, равно как и фарадическая возбудимость мышцы; сокращение же мышцы при раздражении гальваническим током также возникает медленно. Полная реакция перерождения еще не является плохим прогностическим признаком: при условии восстановления (регенерации) нервного волокна она может через фазу частичной реакции замениться нормальной электровозбудимостью. Но если мышца при периферическом параличе остается полностью денервированной свыше 12—14 месяцев (иногда и дольше), то в результате прогрессирующей дегенерации мышечных волокон они погибают полностью, заменяются жировой и соединительной тканью, и наступает цирроз мышцы с утратой уже и реакции ее на гальванический ток, т.е. развивается полная утрата электровозбудимости. Последняя указывает на необратимость происшедших в мышце изменений. Изменения электровозбудимости при периферическом параличе представлены в таблице 3 (по М.И.Аствацатурову). Реакция перерождения наблюдается при атрофиях, которые развиваются в результате поражения периферического двигательного неврона. Другие атрофические процессы в мышцах (артрогенные, от бездеятельности, при заболеваниях самого мышечного аппарата) реакцией дегенерации не сопровождаются. Исследование реакции дегенерации имеет в клинике определенное значение и позволяет проводить дифференциальную диагностику мышечных атрофии различной природы. Кроме того, исследование электровозбудимости дает возможность рано установить диагноз нарушений проводимости нерва, сократительной способности мышц и позволяет судить о динамике процесса, уста- Таблица 4
навливая, например, переход от полной реакции перерождения к частичной в процессе восстановления периферического паралича. Для того, чтобы судить о нормальной электровозбудимости нервов и мышц или установить те или иные отклонения от нормы, необходимо знать средние величины электровозбудимости, полученные в результате исследования большого количества здоровых лиц. В таблице 4 для некоторых нервов и мышц указаны минимальные и максимальные в норме величины гальванической возбудимости; порог раздражения определен в миллиамперах. В процессе изучения электровозбудимости было установлено, что сокращение легче всего получается с определенных участков нервов и мышц, с так называемых двигательных точек, или точек раздражения. Существуют особые схемы с указанием их (рис. 10-14). Реакция перерождения, характерная для периферических параличей, относится к категории качественных изменений электровозбудимости. К этой же категории относятся миотоническая и миастеническая
Рис. 10. Расположение моторных точек на лице. 1 — п. hypoglossus; 2 — quadratus menti; 3 - lev-ator menti; 4 — otbicularis oris; 5 — zygomaticus; 6 - orbicularis palpebrarum; 7 — corrugator supercilii; 8- n. faciails; 9— frontalis; 10 — tem-poralis; // - auricularis posterior; 12 — splenius; 13 - n. accessorius.
Рис. 11. Расположение моторных точек на передней поверхности верхней конечности. / - abductor digiti minimi; 2 - opponens dig-iti minimi; 3 - flexor digiti minimi; 4 - lum-bricales; 5- palmaris brevis; 6- n. ulnaris; 7-flexor digitorum sublimis (IV, V); 8 - flexor digitorum sublimis (II, III); 9 - flexor digitorum communis profundus; 10 — flexor carpiul-naris; 11 - n. ulnaris; 12- triceps (caput inter-num); 13 - triceps (caput longum); 14 - del-loideus; 15 - n. musculocutaneus; 16 - biceps brachii; 77-brachiaIis internus; 18— n. medi-anus; 19 — supinator longus; 20 — pronator teres; 21 - flexor carpi radialis; 22- flexor digitorum sublimis; 23 — flexor pollicis longus; 24 — abductor pollicis brevis; 25 — opponens pollicis; 26 - flexor pollicis brevis; 27 -adductor pollicis. реакции. При миотонии возбудимость нерва остается нормальной, мышца же после полученного сокращения расслабляется крайне медленно. Для миастении характерна крайняя утомляемость мышцы, сказывающаяся в быстром истощении сократительной способности ее при повторных раздражениях током. К количественным изменениям электровозбудимости нервов и мышц относятся: 1) повышение ее, когда для получения сокращения Рис. 12. Расположение моторных точек на задней поверхности верхней конечности. / - inierossei dorsales (I, II); 2 - extensor policis brevis; 3 — abductor pollicis longus; 4 — extensor indicis proprius; 5 — extensor digitorum communis; 6 - extensor carpi radialis brevis; 7 - extensor carpi radialis longus; 8 -supinator longus; 9 - brachialis internus; 10- n. radialis; 11 - deltoideus; 12, 13 - triceps; 14 - extensor caipi ulnaris; 15 - supinator brevis; 16 - extensor digiti minimi; 17 — supinator indicis; 18 - extensor pollicis longus; 19 — abductor digiti minimi; 20- interossei dorsales (III, IV).
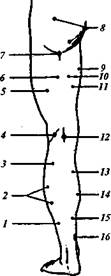
Рис. 13. Расположение моторных точек на передней поверхности нижней конечности. 1 - vastus interims; 2 - cruralis; 3 - adductor longus; 4 — adductor magnus; 5 — pectineus; 6 - obturatorius; 7 - n. femoralis; 8 - tensor fasciae latae; 9 - sartorius; 10 - quadriceps femoris; 11 - rectus femoris; 12 - vastus externus. Рис. 14. Расположение моторных точек на задней поверхности нижней конечности. / - flexor hallucis longus; 2 - soleus; 3 - gas-trocnemius (caput externum); 4 — n. per-oneus; 5 — biceps femoris (caput brevis); 6 — biceps femoris (caput longurn); 7 — n. ischiadicus; 8 — gluteus maximus; 9 — adductor magnus; 10 - semitendinosus; // - semi-mebranosus; 12 - n. tibialis; 13 - gastroc-nemjus (caput internum); 14 — soleus; 75 -flexor digitorum communis longus; 16 — n. tibialis. требуются токи меньшей, чем в норме, силы, или 2) понижение электровозбудимости, когда для получения эффекта необходимо применение токов большей силы, чем у здоровых людей. Другим количественным методом исследования электровозбудимости нервов и мышц является хронаксиметрия. Было установлено, что действие тока определяется не только интенсивностью его, но и длительностью действия на нерв или мышцу. Сначала определяется реобаза, т.е. та минимальная интенсивность постоянного тока, которая необходима, чтобы вызвать эффект - сокращение. Затем применяется ток в два раза большей интенсивности (двойная реобаза) и определяется в тысячных долях секунды (сигмах)
Рис. 15. Нормальная кривая «интенсивность — длительность». На оси ординат показана интенсивность раздражения (в В), на оси абсцисс — его пороговая длительность (в мс). Знак Р -реобаза, знак Хр - хронаксия (при двойной реобазе), см. текст. специальным аппаратом (хронаксиметром) минимальное время, достаточное для сокращения (хронаксия). Хронаксиметрия позволила установить ряд новых и интересных фактов и закономерностей в физиологии и патологии нервной системы. Учение о хронаксии тесно смыкается с понятием о лабильности (Н.Е.Введенский, А.А.Ухтомский). Так, оказалось, что проксимально расположенные мышцы имеют более короткую хронаксию, чем дистальные; мышца и иннервирую-ший ее нерв имеют почти одинаковую хронаксию; мышцы-синергисты имеют одинаковую хронаксию, тогда как мышцы-антагонисты — иную; сгибатели верхних конечностей имеют хронаксию примерно в 2 раза меньшую, чем разгибатели (на нижних конечностях существует обратное соотношение). В норме хронаксия различных мышц составляет от 0,0001 до 0,001 с, при периферических параличах она значительно больше 0,001 с. При центральных параличах (при пирамидном поражении) еще более усиливается расхождение в цифрах хронаксии сгибателей и разгибателей на руках и, наоборот, уменьшается разница цифр на ногах. При экстрапирамидных поражениях эта разница хронаксии уменьшается. Современным и более точным количественным методом исследования электровозбудимости является определение кривой «интенсивность — длительность», которая отражает соотношение между силой (напряжением) тока и длительностью импульса. Первоначально определяется реобаза. В дальнейшем, по мере изменения длительности импульса (от 100 до 0,02 мс), устанавливается интенсивность тока (в В или А), необходимая для получения сокращения мышцы. В норме кривая имеет вид, как на рисунке 15. При частичной и особенно полной денервации кривая рано и резко поднимается кверху; по ходу восстановления проводимости она постепенно возвращается к норме (см. рис. 16). В норме
Рис. 16. Изменение кривой «интенсивность — длительность» в процессе регенерации нерва. Кривая /- при полной денервации мышцы; кривые П-ГУ' - в разные сроки регенерации; кривая V— при полном восстановлении проводимости нерва. кривая показывает возбудимость нерва, а при денервации она отражает возбудимость денервированных мышечных волокон. Можно исследовать проведение возбуждения по нервам. Раздражают током проксимальный, затем дистальный участки нерва и регистрируют на осциллографе вызванные биопотенциалы мышцы. Выясняется латентный период (в мс) ответа мышцы при стимуляции дистального и проксимального отрезков нерва, после чего вычисляется скорость проведения возбуждения по нерву. Для этого необходимо разделить расстояние (в м) между двумя точками стимуляции нерва на время прохождения импульса между этими точками (в с). В норме скорость проведения импульса составляет для локтевого и срединного нервов 50—65 м/с, для малоберцового нерва 40—60 м/с. В диагностике нервно-мышечных заболеваний все большее значение приобретает метод электромиографии, т.е. регистрации изменений электрических потенциалов мышц. Биотоки усиливаются в миллион и более раз, после чего записываются катодными или шлейфными осциллографами в виде кривых - электромиограмм (ЭМГ). Электромиография проводится при различных состояниях мышц: при расслаблении (тонус «покоя»), при рефлекторных изменениях тонуса (во время активного напряжения других мышц, при глубоком вдохе, раздражении рецепторов, эмоциональном напряжении, умственной деятельности), при произвольных напряжениях (сокращениях).
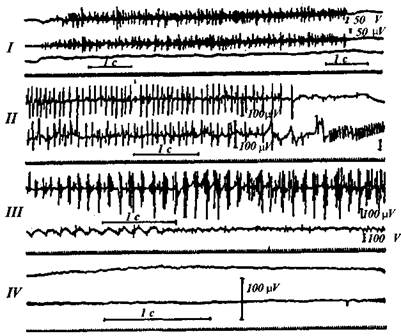
Рис. 17. Основные типы электромиограмм (по классификации Ю.С.Юсевич). /тип — быстрые, частые колебания потенциала, общая структура, амплитуда колебаний меняются в зависимости от формы двигательной реакции и отражают интерференцию электрической активности множества двигательных единиц; // тип — одиночные или ритмически повторяющиеся, редкие, варьирующие по форме колебания потенциала, ЭМГ этого типа отражают электрическую активность отдельных двигательных единиц или синхронизацию колебаний в большем или меньшем их числе; ///тип — сгруппированные в «залпы» частые колебания, иногда сочетающиеся с медленными низковольтными колебаниями, возникающими в ритме «залпов» дрожания; IV тип - полное «биоэлектрическое молчание» — отсутствие колебаний при любых попытках вызвать тонические напряжения или сокращения мышц. Отведение мышечных потенциалов производится с помощью электродов двух видов: поверхностных (накожные диски диаметром около 1 см и межэлектродным расстоянием в 1-2 см) или игольчатых (погружаемых в мышцу). Электроды располагаются в зоне двигательных точек мышц (см. рис. 10-14). Накожные электроды регистрируют суммарную электрическую активность множества мышечных волокон (образцы таких ЭМГ представлены на рисунке 17). Игольчатые электроды с весьма малой отводящей поверхностью и малым межэлектродным расстоянием позволяют изучать электрическую активность немногих или даже отдельных мышечных волокон и мионевральных синапсов (примеры таких записей показаны на рисунке 18).
Рис. 18. Электромиограммы в норме и при поражениях периферического двигательного неврона (по Бухталю). А — здоровая мышца; Б — паралич (полная денервация); В — парез при невральном поражении; Г— парез при поражении клеток переднего рога спинного мозга; /и //- игольчатые электроды и записи от них: а — при расслаблении мышцы (тонус «покоя»); б— при слабом напряжении (повышении тонуса); * - при максимальном произвольном напряжении (сокращении). Объяснение в тексте. Рассмотрим коротко основные варианты ЭМГ в норме, при периферических парезах и некоторых заболеваниях с двигательными расстройствами. На ЭМГ здоровых в покое при локальных отведениях (игольчатыми микро-электродами) колебания потенциала не улавливаются (см. рис. 18, А, а); на суммарной ЭМГ (см. рис. 17, /) можно увидеть очень слабые колебания низкого напряжении. Рефлекторное повышение тонуса сопровождается небольшим усилением электрической активности. Произвольное напряжение здоровых мышц вызывает появление на ЭМГ очень частых высоковольтных колебаний (см. рис. 18, Л, в и рис. 17, Г), при этом общая структура ЭМГ зависит от характера, темпа и силы сокращения. Периферический паралич с полной дегенерацией нервных и мышечных волокон характеризуется исчезновением потенциалов — «биоэлектрическое молчание» (см. рис. 17, IV). Если процесс дегенерации не завершен, то при микроэлектродных отведениях можно отметить слабые и редкие колебания — «потенциалы денервации» (см. рис. 18, Б, а и 6). Парез при поражении нервов или передних корешков вызывает снижение амплитуды колебаний (при произвольном напряжении) в соответствии со степенью нарушения проводимости нерва. При локальных отведениях можно наблюдать потенциалы дегенерации и фибрилляции. Для отличия поражения периферического нерва и переднего корешка необходимо сделать ЭМГ паравертебраль-ных выпрямителей позвоночника, получающих иннервацию от задних ветвей спинальных нервов; эти ветви не входят в состав конечностей. Парез в результате поражения клеток передних рогов (или двигательных ядер черепных нервов) обычно не сопровождается понижением амплитуды колебаний, они могут даже увеличиваться (лишь при грубых парезах амплитуды снижаются), но частота колебаний становится более редкой. Можно уловить потенциалы дегенерации, фибрилляции и фасцикуляции (см. рис. 18, Г). Таким образом, метод электромиографии может помочь в некоторых случаях отличить парезы переднеро-говые от невральных. Мышечные заболевания — миопатии и миозиты — мало отражаются на характере ЭМГ. Амплитуды осцилляции могут понижаться лишь при выраженных атрофиях? Потенциалы денервации отсутствуют. При миотонии и миастении частота и амплитуды биотоков не отличаются от нормы, однако на ЭМГ в первом случае обнаруживается характерная «миотоническая задержка» расслабления, а у больных миастенией нормальная в начале сокращения картина биотоков сменяется прогрессирующим снижением амплитуд. При центральных парезах (пирамидные поражения) во время произвольных сокращений можно наблюдать снижение амплитуд колебаний, но особенно характерны для пирамидных поражений изменения биотоком при рефлекторных повышениях тонуса: синергические напряжения, из
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-20; просмотров: 201; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.188.106.47 (0.018 с.) |