Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Другие органы, вырабатывающие гормоныСодержание книги
Поиск на нашем сайте Желудочно-кишечный тракт вырабатывает большое число различных гормонов (гастрин, мотилин, вазоинтестинальный полипептид, холецистокинин, гастроинтестинальный полипептид и др.), главной функцией которых является регуляция деятельности желудка и кишечника. Плацента образует две группы гормонов - стероидные (прогестерон и эстрогены) и белковые (хорионический гонадотропин -ХГ, плацентарный лактогенныи гормон - ПЛГ и релаксин). Хорионический гонадотропин образуется в больших количествах в первой четверти беременности. Его физиологическая роль сходна с эффектом лютропина передней доли гипофиза. Плацентарный лактогенныи гормон начинает секретироваться у женщин с 6-й недели беременности. Физиологическое значение этого гормона сходно с пролактином передней доли гипофиза. Релаксин усиленно секретируется на поздних стадиях беременности. Он подготавливает организм женщины к родовому акту: расслабляет связки лонного сочленения и других тазовых костей, снижает тонус и сократимость матки. Вилочковая железа - парный дольчатый орган, расположенный в верхнем отделе переднего средостения, образует несколько гормонов: тимозин, тимопоэтин I, тимопоэтин II, тимусный гуморальный фактор, гомеостатический тимусный гормон. Все они являются полипептидами. Гормоны вилочковой железы стимулируют образование антител. Тимус контролирует развитие и распределение лимфоцитов, участвующих в иммунологических реакциях. Недифференцированные стволовые клетки, которые образуются в костном мозге, выходят в кровоток и поступают в ви-лочковую железу, где размножаются и дифференцируются в лимфоциты тимусного происхождения (Т-лимфоциты). Полагают, что именно эти лимфоциты ответственны за развитие клеточного иммунитета. Поступлению гормонов вилочковой железы в кровь способствует соматотропин (гормон роста). Вилочковая железа достигает максимального развития в детском возрасте. После наступления полового созревания она останавливается в развитии и начинает атрофироваться. В связи с этим полагают, что железа стимулирует рост организма и тормозит развитие половой системы. Тимус содержит большое количество витамина С, уступая в этом отношении только надпочечникам. Эпифиз (шишковидная железа) - это образование конусовидной формы, которое нависает над верхними холмиками четверохолмия. По внешнему виду железа напоминает еловую шишку, что и дало повод к ее названию. В эпифизе образуется гормон мелатонин, участвующий в регуляции пигментного обмена (он обесцвечивает меланофоры). Мелатонин тормозит развитие половых функций у молодого организма и угнетает действие гонадотропных гормонов у взрослого, что реализуется посредством гипоталамуса и гипофиза. Глава 5 РЕГУЛЯЦИЯ ДВИЖЕНИЙ Любой двигательный акт человека, как и всех животных, осуществляется благодаря сократительной деятельности скелетных мышц. Мышцы же могут сокращаться только при поступлении к ним нервных импульсов'из центральной нервной системы (ЦНС). Поэтому в данной главе рассмотрим физиологию мышц и ЦНС. СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА СКЕЛЕТНОЙ МЫШЦЫ И МЕХАНИЗМ ЕЕ СОКРАЩЕНИЯ Структурной и функциональной единицей скелетной мышцы является мышечное волокно, представляющее собой сильно вытянутую многоядерную клетку. Длина мышечного волокна зависит от размеров мышцы и составляет от нескольких милли-
метров до нескольких сантиметров. Толщина волокна вариабельна (10-100 мкм). Скелетная мышца - это система, преобразующая химическую энергию в механическую работу и тепло. Специфическими элементами мышечного волокна являются специализированный сократительный аппарат - миофибриллы, системы продольных трубочек - саркоплазматическая сеть (ретикулум) и система поперечных трубочек - Т-система, представляющая собой впячивания поверхностной мембраны мышечного волокна внутрь его. Миофибрилла состоит из нескольких саркомеров, являющихся функциональной единицей сократительного аппарата мышечного волокна. Саркомеры отделяются друг от друга 2-пластинками. Скелетная мышца имеет поперечную исчерченность, которая обусловлена особым расположением сократительных миофиб-рилл - белков актина и миозина: их концы не полностью перекрывают друг друга, что и определяет исчерченность (рис. 5.1). Акти-новые филаменты представлены двойной нитью, закрученной в двойную спираль; они одним концом прикреплены к 2-пластинке, другим концом взаимодействуют с нитями миозина при сокращении мышцы. Нити миозина располагаются в середине саркомера и своими концами взаимодействуют с нитями актина. В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина, к молекуле которого прикреплена молекула другого белка - тропонина. В состоянии покоя нити актина и миозина незначительно перекрывают друг друга таким образом, что общая длина саркомера составляет около 2,5 мкм. На боковых сторонах миозиновой нити имеются выступы, получившие название поперечных мостиков, состоящих из головки и шейки. Головка приобретает выраженную АТФ-азную активность при контакте с актином. При сокращении происходит укорочение саркомера вследствие активного перемещения тонких актиновых нитей относительно толстых миозиновых. Механизм мышечного сокращения. Мышца сокращается в естественных условиях только при поступлении к ней нервных импульсов. Нервное влияние на мышечное волокно передается с помощью нервно-мышечного синапса. Медиатором в нервно-мышечном синапсе является ацетилхолин. На один ПД из пресинаптиче-ского окончания нервно-мышечного синапса выделяется 200-300 квантов медиатора. В состоянии покоя мышцы, т.е. в промежутках между передачей нервного импульса, происходит спонтанное выделение 1-2 квантов медиатора в синаптическую щель в среднем 1 раз в секунду. При этом на постсинаптической мембране формируется деполяризация с амплитудой 0,12-0,24 мВ. Такие потенциалы получили название миниатюрные потенциалы концевой пластинки. Они, вероятно, поддерживают высокую возбудимость синапсов в условиях функционального покоя нервных центров. Кроме эк-зоцитоза медиатора существует постоянная неквантовая утечка молекул медиатора в синаптическую щель. Предполагают, что неквантовая секреция играет трофическую роль. Пришедший по нервному волокну импульс (ПД) обеспечивает выделение в синаптическую щель ацетилхолина, который на пост-• синаптической мембране (концевой пластинке мышечного волокна) вызывает возникновение потенциала концевой пластинки (ПКП) -возбуждающего постсинаптического потенциала (ВПСП), амплитуда которого составляет 30-40 мВ. ПКП - это локальный потенциал, который, достигнув критической величины, обеспечивает возникновение ПД в мышечном волокне. ПД распространяется по мышечному волокну и Т-системе в глубь волокна, что обеспечивает выделение ионов Са2+ из саркоплазматического ретикулума. При взаимодействии ионов Са2+ с тропонином происходят смещение тропомиозина и освобождение активных центров на актиновых нитях. Затем происходит присоединение головки поперечного мостика миозина к актиновой нити. При этом головка миозина приобретает АТФ-азную активность, что обеспечивает гидролиз АТФ и освобождение энергии, обеспечивающей поворот головки миозина вокруг своей оси (гребковое действие), что в свою очередь приво- дит к скольжению нитей актина и миозина относительно друг друга и укорочению саркомера и общей длины мышцы. Миозиновая головка имеет несколько активных центров, которые последовательно взаимодействуют с соответствующими центрами на актиновой нити. В каждый конкретный момент в процессе развития сокращения одни головки поперечных мостиков находятся в соединении с актиновой нитью, другие свободны, т. е. существует последовательность их взаимодействия с актиновой нитью, что обеспечивает плавность процесса сокращения. Повторное присоединение миозиновой головки к новому центру на актиновой нити вновь приводит к повороту головки, который обеспечивается запасенной в ней энергией. В каждом цикле соединения и разъединения головки миозина с актином расщепляется одна молекула АТФ на каждый мостик. Быстрота поворота определяется скоростью расщепления АТФ. Для расслабления мышцы в первую очередь необходимо понижение концентрации ионов Са2+ в области сократительных элементов мышечного волокна. Саркоплазматическая сеть имеет кальциевый насос, который активно возвращает кальций в цистерны. Активация кальциевого насоса осуществляется неорганическим фосфатом, который образуется при гидролизе АТФ, а энергообеспечение работы кальциевого насоса также осуществляется за сче* энергии, образующейся при гидролизе АТФ. Источником энергии для восстановления израсходованной АТФ являются белки, жиры и углеводы пищи, которые подвергаются расщеплению в желудочно-кишечном тракте и в виде мономеров поступают в кровь и лимфу. В организме в результате биохимических превращений образуется АТФ или синтезируются крупномолекулярные вещества. АТФ - донор свободной энергии в клетках. В клетках АТФ используется в течение одной минуты после ее образования, скорость оборота АТФ очень велика. Стабильность концентрации АТФ в клетке поддерживается рядом механизмов, одним из которых является образование креатинфосфата (КФ). Когда количество АТФ превышает определенный уровень, часть ее энергии используется для синтеза КФ, количество которого при этом возрастает. При повышении же распада АТФ в условиях активации энергетического обмена КФ используется для ресинтеза АТФ с помощью окисления и фосфорилирования. Существуют и бескислородные (анаэробные) пути преобразования энергии, в которых могут быть использованы только углеводы (анаэробный гликолиз); такие способы реализуются при недостаточном поступлении кислорода в организм, ткани и клетки. При полном прекращении дыхания и расходовании резервов кислорода эти процессы могут обеспечить потребность в энергии еще в течение двух минут. Запас АТФ в скелетных мышцах обеспечивает всего лишь 10 одиночных сокращений. При максимальном мышечном сокращении имеющихся в тканях запасов АТФ достаточно лишь на одну секунду. Энергия КФ, концентрация которого в три-восемь раз больше, чем АТФ, может поддержать такое сокращение в течение еще нескольких секунд. При максимальном сокращении на протяжении нескольких секунд абсолютно необходим анаэробный гликолиз, в котором используются запасы гликогена. Ресинтез гликогена из образующейся при этом молочной кислоты возможен, однако, лишь в аэробных условиях. Аэробное окисление глюкозы и жирных кислот в цикле Кребса, совершаемое в митохондриях, - это наиболее типичный способ энергообеспечения скелетных мышц. Запасы свободной глюкозы, гликогена и жиров в мышцах достаточно велики. Однако при длительной работе в организме накапливаются недоокисленные продукты (молочная кислота и др.). Создается кислородная задолженность. Такой долг погашается после работы за счет компенсаторной мобилизации кровообращения и дыхания (тахикардия, повышение кровяного давления, одышка). Если же работа, несмотря на наличие кислородного долга, продолжается, наступает выраженное состояние утомления, которое иногда прекращается за счет мобилизации дополнительных резервов кровообращения и дыхания («второе дыхание» спортсменов). Таким образом, энергия АТФ в скелетной мышце используется для трех процессов: 1) работы № / К-насоса, обеспечивающего поддержание постоянства градиента концентрации этих ионов по обе стороны мембраны; 2) процесса скольжения актиновых и миозино-вых нитей, приводящих к укорочению миофибрилл (комплекс актин - миозин становится стабильным только при трупном окоченении, когда концентрация АТФ в мышце падает ниже некоторой критической величины)* 3) работы Са-насоса, активируемого при расслаблении волокна. ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ В зависимости от изменения длины мышечного волокна выделяют два типа его сокращения - изометрическое и изотоническое. Сокращение мышцы, при котором ее волокна укорачиваются при неизменном напряжении, называется изотоническим. Сокра-
щение мышцы, при котором ее напряжение возрастает, а длина мышечных волокон остается неизменной, называется изометрическим. В естественных условиях сокращения мышц являются смешанными - мышца обычно не только укорачивается, но изменяется и ее напряжение. В зависимости от длительности выделяют одиночное и тетаническое сокращения мышцы. Одиночное сокращение мышцы в эксперименте вызывают одиночным раздражением электрическим током мышцы или нервного волокна. В изотоническом режиме одиночное сокращение начинается через короткий скрытый (латентный) период, далее следует фаза подъема (фаза укорочения), затем фаза спада (фаза расслабления) (рис. 5.2). Обычно мышца укорачивается на 5-10% исходной длины. Длительность ПД мышечных волокон также варьирует и составляет 5-10 мс с учетом замедления фазы реполяризации в конце ее. Длительность одиночного сокращения, мышечного волокна вариабельна, она во много раз превышает продолжительность ПД. Мышечное волокно подчиняется закону «все или ничего», т.е. отвечает на пороговое и сверхпороговое раздражение одинаковым по величине одиночным сокращением. Однако сокращение целой мышцы зависит от силы раздражения при непосредственном раздражении мышцы и от числа нервных импульсов, поступающих к мышце при раздражении нерва. Пр'и непосредственном раздражении это связано с различной возбудимостью мышечных волокон и разным расстоянием их от раздражающих электродов. Увеличение силы раздражения ведет к увеличению числа сокращающихся мышечных волокон. Подобный эффект наблюдается и в естественных условиях -с увеличением числа возбужденных нервных волокон и частоты импульсов (к мышце поступает больше нервных импульсов - ПД) увеличивается число сокращающихся мышечных волокон. При одиночных сокращениях мышца утомляется незначительно. Тетаническое сокращение - это слитное длительное сокращение скелетной мышцы. В его основе лежит явление суммации одиночных мышечных сокращений. При нанесении на мышечное
волокно или непосредственно на мышцу двух быстро следующих друг за другом раздражений возникающее сокращение имеет большую амплитуду и длительность. При этом нити актина и миозина дополнительно скользят друг относительно друга. Могут вовлекаться также в сокращение ранее не сокращавшиеся мышечные волокна, если первый стимул вызвал у них подпороговую деполяризацию, а второй увеличивает ее до критической величины. Суммация сокращений при повторном раздражении мышцы или поступлении к ней ПД возникает только в том случае, когда закончен рефрактерный период (после исчезновения ПД мышечного волокна). При поступлении импульсов к мышце во время ее расслабления возникает зубчатый тетанус, во время укорочения - гладкий тетанус (рис. 5.3). Амплитуда тетануса больше величины максимального одиночного сокращения мышцы. Напряжение, развиваемое мышечными волокнами при гладком тетанусе, обычно в 2-4 раза больше, чем при одиночном сокращении, однако мышца быстрее утомляется. Мышечные волокна не успевают восстановить энергетические ресурсы, израсходованные во время сокращения. Амплитуда гладкого тетануса увеличивается с возрастанием частоты стимуляции нерва. При некоторой {оптимальной) частоте стимуляции амплитуда гладкого тетануса наибольшая (опти-
мум частоты раздражения). При чрезмерно частой стимуляции нерва (более 100 имп/с) мышца расслабляется вследствие блока проведения возбуждения в нервно-мышечных синапсах - песси-мум Введенского (пессимум частоты раздражения). Пессимум Введенского можно получить и при прямом, но более частом раздражении мышцы (более 200 имп/с) (см. рис. 5.3). Пессимум Введенского не является результатом утомления мышцы или истощения медиатора в синапсе, что доказывается фактом возобновления сокращения мышцы сразу же после уменьшения частоты раздражения. Торможение развивается в нервно-мышечном синапсе при раздражении нерва. В естественных условиях мышечные волокна сокращаются в режиме зубчатого тетануса или даже одиночных последовательных сокращений. Однако форма сокращения мышцы в целом напоминает гладкий тетанус. Причины этого - асинхронность разрядов мотонейронов и асинхронность сократительной реакции отдельных мышечных волокон, вовлечение в сокращение большого их количества, вследствие чего мышца плавно сокращается и плавно расслабляется, может длительно находиться в сокращенном состоянии за счет чередования сокращений множества мышечных волокон. При этом мышечные волокна каждой двигательной единицы сокращаются синхронно. ДВИГАТЕЛЬНЫЕ ЕДИНИЦЫ Понятия. Иннервация скелетных мышечных волокон осуществляется мотонейронами спинного мозга или мозгового ствола. Один мотонейрон веточками своего аксона иннервирует несколько мышечных волокон. Совокупность мотонейрона и иннервируемых им мышечных волокон называют двигательной (нейромоторной) единицей. Число мышечных волокон двигательной единицы варьирует в широких пределах в разных мышцах. Двигательные единицы невелики в мышцах, приспособленных для быстрых движений, от нескольких мышечных волокон до нескольких десятков их (мышцы пальцев, глаза, языка). Наоборот, в мышцах, осуществляющих медленные движения (поддержаниеттозы мышцами туловища), двигательные единицы велики и включают сотни и тысячи мышечных волокон. При сокращении мышцы в натуральных (естественных) условиях можно зарегистрировать ее электрическую активность (элек-тромиограмму - ЭМГ) с помощью игольчатых или накожных электродов. В абсолютно расслабленной мышце электрическая активность почти отсутствует. При небольшом напряжении, например при поддержании позы, двигательные единицы разряжаются с небольшой частотой (5-10 имп/с), при большом напряжении частота импульсации повышается в среднем до 20-30 имп/с. ЭМГ позволяет судить о функциональной способности нейромоторных единиц. С функциональной точки зрения двигательные единицы разделяют на медленные и быстрые. Медленные двигательные единицы включают медленные мотонейроны и медленные мышечные волокна (красные). Медленные мотонейроны, как правило, низкопороговые, так как обычно это малые мотонейроны. Устойчивый уровень импульсации у медленных мотонейронов наблюдается уже при очень слабых статических сокращениях мышц, при поддержании позы. Медленные мотонейроны способны поддерживать длительный разряд без заметного снижения частоты импульсации на протяжении длительного времени. Поэтому их называют малоутомляемыми или не-утомляемыми мотонейронами. В окружении медленных мышечных волокон богатая капиллярная сеть, позволяющая получать большое количество кислорода из крови. Повышенное содержание миоглобина облегчает транспорт кислорода в мышечных клетках к митохондриям. Миоглобин обусловливает красный цвет этих волокон. Кроме того, волокна содержат большое количество митохондрий и субстратов окисления - жиров. Все это обусловливает использование медленными мышечными волокнами более эффективного аэробного окислительного пути энергопродукции и определяет их высокую выносливость. Быстрые двигательные единицы состоят из быстрых мотонейронов и быстрых мышечных волокон. Быстрые высокопороговые мотонейроны включаются в активность только для обеспечения относительно больших по силе статических и динамических сокращений мышц, а также в начале любых сокращений, чтобы увеличить скорость нарастания напряжения мышцы или сообщить движущейся части тела необходимое ускорение. Чем больше скорость и сила движений, т. е. чем больше мощность сократительного акта, тем больше участие быстрых двигательных единиц. Быстрые мотонейроны относятся к утомляемым - они не способны к длительному поддержанию высокочастотного разряда. Быстрые мышечные волокна (белые мышечные волокна) более толстые, содержат больше миофибрилл, обладают большей силой, чем медленные волокна. Эти волокна окружает меньше капилляров, в клетках меньше митохондрий, миоглобина и жиров. Активность окислительных ферментов в быстрых волокнах ниже, чем в медленных, однако активность гликолитических ферментов, запа-
сы гликогена выше. Эти волокна не обладают большой выносливостью и более приспособлены для мощных, но относительно кратковременных сокращений. Активность быстрых волокон имеет значение для выполнения кратковременной высокоинтенсивной работы, например бега на короткие дистанции. Выделяют также тонические мышечные волокна, они имеют 7-10 синапсов, принадлежащих, как правило, нескольким мотонейронам. ПКП этих мышечных волокон не вызывает генерацию ПД в них, а непосредственно запускает мышечное сокращение. Скорость сокращения мышечных волокон находится в прямой зависимости от активности миозин-АТФ-азы - фермента, расщепляющего АТФ и тем самым способствующего образованию поперечных мостиков и взаимодействию актиновых и миозиновых мио-филаментов. Более высокая активность этого фермента в быстрых мышечных волокнах обеспечивает и более высокую скорость их сокращения по сравнению с медленными волокнами. СИЛА МЫШЦЫ И ЕЕ РАБОТА В различных мышцах тела соотношение между числом медлен-,, ных и быстрых мышечных волокон неодинаково, поэтому и сила их сокращения, и степень укорочения вариабельны. При снижении физической нагрузки - особенно большой интенсивности, при которой требуется активное участие быстрых мышечных волокон, - последние истончаются (гипотрофируются) быстрее, чем медленные волокна, быстрее уменьшается их число. А. Факторы, влияющие на силу сокращения мышцы. 1. Число сокращающихся волокон в данной мышце. С увеличением сокращающихся волокон возрастает сила сокращений мышцы в целом. В естественных условиях сила сокращения мышцы возрастает с увеличением нервных импульсов, поступающих к мышце, в эксперименте - с увеличением силы раздражения. 2. Соотношение быстрых и медленных волокон. Чем больше быстрых волокон содержит мышца, тем больше возможная ее сила сокращения. 3. Поперечное сечение мышцы. Различают геометрическое и физиологическое поперечные сечения мышцы. Геометрическое поперечное сечение перпендикулярно продольной оси мышцы, физиологическое - длине мышечных волокон. В портняжной мышце, например, все волокна параллельны длине оси мышцы - парал-лельноволокнистый тип. Большинство мышц - перистого ' 84 типа, их волокна расположены косо, прикрепляясь с одной стороны к центральному сухожильному тяжу, а с другой стороны - к наружному сухожильному футляру. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косым расположением волокон физиологическое поперечное сечение может значительно превосходить геометрическое поперечное сечение. Термины «абсолютная» и «относительная» сила мышцы нередко путаются, так как в них заложена идея, противоположная общепринятой в подобных случаях, поэтому необходимо пользоваться другими: «общая сила мышцы» (определяется максимальным напряжением в кг, которое она может развить) и «удельная сила мышцы» - отношение этого напряжения в кг к физиологическому поперечному сечению мышцы (кг/см2). Чем больше физиологическое поперечное сечение мышцы, тем больший груз она в состоянии поднять. По этой причине сила мышцы с косо расположенными волокнами больше силы, развиваемой мышцей той же толщины, но с продольным расположением волокон. Для сравнения силы разных мышц максимальный груз, который они в состоянии поднять, делят на площадь их физиологического поперечного сечения (удельная сила мышцы). Вычисленная таким образом сила (кг/см2) для трехглавой мышцы плеча человека - 16,8, двуглавой мышцы плеча - 11,4, сгибателя плеча - 8,1, икроножной мышцы - 5,9, гладких мышц - 1 кг/см2.
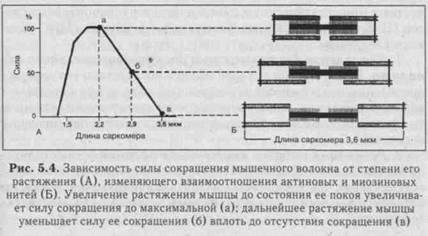
4. Умеренное растяжение мышцы также ведет к увеличению ее сократительного эффекта. Однако при чрезмерном растяжении сила сокращения уменьшается. Это демонстрируется в опыте с дозированным растяжением мышцы (рис 5.4). Если мышца перерас- тянута так, что нити актина и миозина не перекрываются, то общая сила мышцы равна нулю. По мере приближения к натуральной длине покоя, при которой все головки миозиновых нитей способны контактировать с актиновыми нитями, сила мышечного сокращения вырастает до максимума. Однако при дальнейшем уменьшении длины мышечных волокон из-за перекрытия нитей актина и миозина сила сокращения мышцы снова уменьшается вследствие уменьшения возможной зоны контакта нитей актина и миозина. Подсчитано, что одиночное мышечное волокно способно развить напряжение 100-200 мг. 5. Функциональное состояние мышцы. При утомлении мышцы величина ее сокращения снижается. Б. Работа мышцы измеряется произведением поднятого груза на величину ее укорочения. Зависимость мышечной работы От нагрузки подчиняется закону средних нагрузок. Если мышца сокращается без нагрузки, ее внешняя работа равна нулю. По мере увеличения груза работа увеличивается, достигая максимума при средних нагрузках. Затем она постепенно уменьшается с увеличением нагрузки. Работа становится равной нулю при очень большом грузе, который мышца при своем сокращении не способна поднять.
|
||||||
|
|
Последнее изменение этой страницы: 2016-04-18; просмотров: 490; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.034 с.) |