
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Структура и функция сетчаткиСодержание книги
Поиск на нашем сайте
Сетчатка у млекопитающих животных состоит из нескольких слоев клеток различного типа. Прежде чем достигнуть фоторецеп-торных клеток, световые лучи должны пройти через все слои этих клеток (см. рис. 15.16, Б). Фоторецепторные клетки — это сильно видоизмененные волосковые клетки. Сома клетки — внутренний сегмент содержит ядро и большое количество мелких митохондрий (рис. 15.17, А); в базальной части имеется афферентное синаптичес-кое окончание, а от верхнего конца отходит рудиментарная реснич- 43* 675
Г-- Канал открывается Снижение натриевой проводимости Синоптическое окончание
cNa Потенциал поко
я Синоптическое окончание Na Свет
Морфофункциональная организация фоторецепторов позвоночных животных:
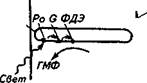
А. Палочка и колбочка сетчатки. Б. Схема передачи фотовозбуждения к ионным каналам наружной мембраны рецептора: Р. — фотопигмент родопсин; С —белок; ФДЭ — фосфодиэсте-раза; ГМФ — гуанозинмонофосфат; цГМФ — циклический гуанозинмонофосфат. В. Изменение ионного тока через мембрану фоторецептора при освещении. Г. Изменение мембранного потенциала (потенциала покоя) фоторецептора при освещении: £ма — равновесный потенциал для ионов натрия; Ек — равновесный потенциал для ионов калия. По оси ординат — время
ка, внутри которой, как и в киноцилии, по кругу располагаются 9 пар фибрилл. Ресничка в одних случаях образует расширение — наружный сегмент в виде цилиндра, в других — в виде колбы. Соответственно первый тип фоторецептора получил название палочки, а второй — колбочки. У колбочек мембрана образует дис-
кообразные, накладывающиеся друг на друга ш.шичшытш уменьшающиеся в диаметре в направлении к верхнему и»ин\ внутренняя полость каждого диска сообщается с внутри и ми средой колбочки. У палочек мембраны дисков полностью зам кнуты и внутреннее пространство диска изолировано от внутренней среды наружного сегмента палочки. Диски собраны стопкой и заключены в футляр из поверхностной мембраны наружного сегмента. В мембраны дисков обоих типов фоторецепторов встроен интегральный фоторецепторный белок — фотопигмент, концентрация которого весьма значительна. Например, у коров на долю фотопигмента приходится около 80 % общего количества белков мембраны. Все это указывает на то, что процессы фотохимического преобразования энергии светового стимула происходят именно в дисках. Большое количество дисков увеличивает светочувствительную поверхность фоторецептора. Зрительные диски образуются на протяжении всей жизни животного. Они постепенно в течение нескольких недель перемещаются по наружному сегменту с последующим отделением от него, поглощением и разрушением клетками пигментного эпителия. Пигментный эпителий выполняет еще ряд важных функций: препятствует отражению и рассеиванию световых лучей, способствует более четкому зрительному восприятию. Наружные сегменты фоторецепторов окружены отростками пигментного эпителия. При сильном освещении частицы пигмента перемещаются из эпителиальных клеток и экранируют наружные сегменты от яркого света. При регистрации электрической реакции фоторецепторов на световой стимул было обнаружено, что палочки и колбочки генерируют только гиперполяризационные рецепторные потенциалы (см. рис. 15.17, Г). Измерение проводимости до освещения и во время действия светового стимула показало, что в темноте мембрана наружного сегмента примерно в одинаковой мере проницаема для ионов натрия и калия. Поэтому (см. гл. 2) согласно уравнению Голдмана величина покоя фоторецептор-ных клеток находится приблизительно между равновесным потенциалом для ионов калия и натрия. Ионы натрия входят в наружный сегмент по своему градиенту через каналы, которые в темноте открыты. Важно отметить, что при генерации темнового тока ионы натрия не накапливаются во внутриклеточном пространстве благодаря интенсивному функционированию Na+ — К+-ионного насоса. Не случайно в фоторецепторах сосредоточено во внутреннем сегменте большое количество митохондрий. При освещении натриевые каналы закрываются, что приводит к уменьшению темнового тока и к сдвигу мембранного потенциала в сторону равновесного калиевого потенциала, т. е. к гиперполяризации мембраны. После окончания действия светового стимула мембранный потен-
циал вновь сдвигается в положительном направлении и занимает уровень между равновесными калиевым и натриевым потенциалами. Изменения мембранного потенциала передаются путем простого кабельного распространения тока на базальную часть внутреннего сегмента клетки, где модулируют процесс высвобождения нейромедиатора из пресинаптического участка клетки. Разберем механизм передачи возбуждения от фоторецепторно-го белка к ионным каналам. Согласно электрофизиологическим, биохимическим и биофизическим исследованиям фотоактивируе-мый белок и ионные каналы в фоторецепторах пространственно разнесены: внутриклеточным посредником выступают молекулы циклического гуанозин-3', 5'-монофосфата (цГМФ). В темноте цГМФ «удерживает» ионные натриевые каналы в открытом состоянии. Поглощение света фотопигментом (родопсином) вызывает активацию G-белка. Активированный G-белок, в свою очередь, активирует фермент фосфодиэстеразу, которая гидролизует цГМФ, превращая его в обычный ГМФ. Снижение концентрации цГМФ приводит к закрытию натриевых каналов и темновой ток уменьшается. После светового стимула в темноте под действием другого фермента — гуанилатциклазы происходит восстановление концентрации цГМФ. Повышение уровня цГМФ ведет к открытию натриевых каналов и восстановлению темнового тока до первоначального уровня. То, что цГМФ является вторичным посредником в фотоэлектрических процессах, позволяет объяснить чрезвычайно высокую чувствительность фоторецепторов. Так, поглощение рецепторной клеткой нескольких фотонов, энергия которых ничтожна, вызывает заметную электрическую реакцию фоторецеп-торной клетки, превышающей в десятки тысяч раз первоначальную энергию фотостимула. Примерная, в определенной степени заниженная оценка показывает, что фотовозбуждение одной молекулы фотопигмента активирует 10 молекул G-белка. Далее одна молекула G-белка активирует 10 молекул фосфодиэстеразы, которая, в свою очередь, гидролизует 10 молекул цГМФ. Таким образом активация 1 молекулы фотобелка уменьшает содержание цГМФ на 1000 молекул, т. е. происходит усиление реакции в 1000 раз. Большие успехи достигнуты в изучении структуры и фотохимических превращений фотопигментов в фоторецепторных клетках при нанесении светового стимула. Предположение о том, что для процесса фоторецепции нужен какой-то пигмент, было выдвинуто в конце XVIII в. Д. Дрейпером. Вскоре после этого В. Кюне выделил чувствительное к свету вещество — родопсин, которое обусловливает пурпурный цвет сетчатки. Им же было показано, что после обесцвечивания пигмента на свету (пурпурный цвет бледнеет) его цвет восстанавливается через некоторое время в темноте при условии сохранения контакта между рецепторными клетками и пигментным эпителием. Родопсин содержится в палочках всех млекопитающих. Плотность молекул родопсина в фоторецепторной мембране весьма велика — 5- III1' мпнгкуи на 1 см2 мембраны. Родопсин лучше всего поглощаем i шч < пни ной волны около 500 нм (сине-зеленая область). В шим.ниннгм были определены другие пигменты. Все изученные фотопигменты состоят из двух глаииыч номмп нентов: белка опсина и простетической группы, которая и шиш* случаях представлена ретиналем (альдегидная форма витамина А|), в других — 3-гидроретиналем (альдегидная форма витамина А;) Действие света на родопсин (а также на другие зрительные иш менты) можно представить в виде следующей общей схемы: Свет Родопсин------- > Ретиналь + Опсин <------------ Темнота ↕ Ретинол В темноте опсин и ретиналь тесно связаны друг с другом. Как предполагают, ретиналь точно «входит» как ключ в замок в определенный участок молекулы опсина. Поглощение светового кванта вызывает цепь конформационных превращений и в ретинале, и в опсине. Родопсин, в котором ретиналь находится в форме изогнутой молекулы — в г^ыс-форме, превращается в люмиродопсин — неустойчивое соединение с ретиналем в транс-форме, с выпрямленной боковой цепью. Необходимо отметить, что цис-транс-vao- меризация является единственным эффектом, вызываемым светом в зрительном пигменте. Все последующие реакции происходят спонтанно с выделением энергии при физиологических температурах (т. е. при обычной температуре тела животного). Люмиродопсин превращается в метародопсин. Измененная форма опсина, очевидно, вызывает активацию G-белка, запускающего ферментативный каскад реакций, в результате чего происходит гидролиз цГМФ и закрытие натриевых каналов. Метародопсин спонтанно гидролизуется до ретиналя и опсина. 7)?анс-форма ретиналя переходит в ретинол (витамин АО, т. е. вновь идет процесс изомеризации — превращение в г<«с-форму. После этого г<ис-ретиналь с белком опсином в темноте формируют родопсин, который вновь участвует в фотокинетическом процессе. При очень ярком свете происходит интенсивное обесцвечивание родопсина. Для достижения исходного уровня родопсина требуется некоторое время, и чувствительность фоторецепторов в этот период снижена. Если в это время животное или человека поместить в темное помещение, то окружающие предметы мало различимы. Однако по мере восстановления уровня родопсина чувстви-
ЦВЕТОВОЕ ЗРЕНИЕ Основы теории цветового зрения были заложены художниками эпохи Возрождения. Они хорошо знали, что можно воспроизвести любой цвет, комбинируя три основные краски: красную, желтую и синюю. На основании этих данных М. В. Ломоносов предположил, что в глазу имеются структуры, обеспечивающие цветное восприятие путем комбинации трех цветов. Им впервые в 1751 г. была выдвинута идея о «трех материях дна ока». Подобная идея была высказана спустя 50 лет Т. Юнгом, который предположил, что рецепторы глаза избирательно воспринимают три основных цвета: красный, желтый и синий. Кроме того, рецепторы каждого типа в меньшей мере могут возбуждаться и другими цветами. Например, «красные» и «желтые» рецепторы наряду с тем, что будут давать максимальную реакцию на соответствующие основные цвета," будут реагировать и на оранжевый. Таким образом, согласно Т. Юнгу, ощущение «оранжевого цвета» возникает в результате одновременного возбуждения «красных» и «желтых» рецепторов. Трихроматические теории Ломоносова — Юнга подтвердили в XIX—XX вв. многочисленные психофизические эксперименты Максвела, Гельмгольца и Рэштона. Однако объективные доказательства о существовании в сетчатке глаза трех типов цветных рецепторов были получены впервые в 60-х годах XX в. У. Марксом и Э. МакНиколом. Изучая спектры поглощения одиночных колбочек из сетчатки золотой рыбки, они обнаружили три типа колбочек, которые различались по спектральным пикам поглощения световых волн и соответствовали трем зрительным пигментам. Подобного типа исследования, проведенные на сетчатке млекопитающих, дали сходные результаты. В соответствии с фотохимическими законами свет, состоящий из волн разной длины, стимулирует фотохимические реакции пропорционально поглощению световых волн каждой длины (каждого цвета). В том случае, если фотон не поглощается, то никакого влияния на молекулу пигмента он не оказывает. I loi лощенный же фотон передает часть своей энергии молекуле пигмента и hi.i ii.iiui ет цепь реакций, в результате которых клетка генерирует рент торный потенциал (см. рис. 15.17, Б). Таким образом, полны p.i i ной длины (разного цвета) будут возбуждать фоторецепторпук» клетку пропорционально тому, насколько эффективно пигмеиi данной клетки поглощает эти волны, т. е. в соответствии с ее спектром поглощения света. Цветовое зрение было выявлено у представителей всех клас сов позвоночных. Вместе с тем к настоящему времени экспериментальные данные не позволяют сделать окончательные выводы о вкладе палочек и колбочек в восприятие цвета. Как правило, цветовое зрение связано с присутствием в сетчатке колбочек, однако в ряде случаев были обнаружены и «цветные» типы палочек. Морфологические отличия между тремя видами колбочек не выяснены. Поскольку структура и свойства ретиналя остаются во всех фоторецепторах неизменными, можно считать, что цветочувствительность разных фотопигментов связана с изменениями структуры опсина. Родопсины человека имеют максимумы чувствительности в синей, зеленой и желтой частях спектра. Спектральные характеристики глаз животных и человека отличаются. Так, лошади, овцы и свиньи различают лишь красные и зеленые цвета. Цветовое ощущение, возникающее в ЦНС у человека и животных, очевидно, определяется соотношением между электрическими сигналами на выходе колбочек того или иного типа.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2016-12-16; просмотров: 281; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.237.229 (0.013 с.) |

 Наружная мембрана
Наружная мембрана цГМФ^^р закрывается
цГМФ^^р закрывается Темнота В
Темнота В тельность фоторецепторов к свету возрастает и глаза начинают различать окружающую обстановку. Этот процесс приспособления называют темповой адаптацией. В норме содержание рети-наля в фоторецепторах в результате фотохимических процессов восполняется из витамина А{ (ретинола), запасенного в клетках пигментного эпителия, в которые он поступает из крови. Недостаток витамина Ах в пище приводит к замедлению образования ретиналя и соответственно родопсина. Это сопровождается заметным снижением чувствительности глаза к свету. Глаз теряет способность к темновой адаптации, развивается так называемая «куриная слепота».
тельность фоторецепторов к свету возрастает и глаза начинают различать окружающую обстановку. Этот процесс приспособления называют темповой адаптацией. В норме содержание рети-наля в фоторецепторах в результате фотохимических процессов восполняется из витамина А{ (ретинола), запасенного в клетках пигментного эпителия, в которые он поступает из крови. Недостаток витамина Ах в пище приводит к замедлению образования ретиналя и соответственно родопсина. Это сопровождается заметным снижением чувствительности глаза к свету. Глаз теряет способность к темновой адаптации, развивается так называемая «куриная слепота».


