
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Транспорт газов кровью, газообмен в тканяхСодержание книги
Поиск на нашем сайте
Транспорт кислорода кровью. Кислород транспортируется кровью в двух формах — в растворенном виде и в соединении с гемоглобином. В плазме артериальной крови содержится очень небольшое количество физически растворенного кислорода, всего 0,3 об.%, т. е. 0,3 мл кислорода в 100 мл крови. Основная же часть кислорода вступает в непрочное соединение с гемоглобином эритроцитов, образуя оксигемоглобин. Насыщение крови кислородом называется оксигенацией или артериализацией крови. Кровь, оттекающая от легких по легочным венам, имеет такой же газовый состав, что и артериальная кровь в большом круге кровообращения. Количество кислорода, находящееся в 100 мл крови при условии полного перехода гемоглобина в оксигемоглобин, называется кислородной емкостью крови. Эта величина помимо парциального давления кислорода зависит от содержания гемоглобина в крови. Известно, что 1 г гемоглобина может в среднем связать 1,34 мл кислорода. Следовательно, зная уровень содержания в крови гемоглобина, можно вычислить кислородную емкость крови. Так, у лошадей при содержании гемоглобина в крови около 14 г/100 мл кислородная емкость крови составляет (1,34 • 14) около 19 об.%, у крупного рогатого скота при уровне гемоглобина 10... 12 г/100 мл — около 13...16 об.%. Пересчитав содержание кислорода в общем объеме крови, оказывается, что его запаса хватит лишь на З...4мин при условии, если он не будет поступать из воздуха. На уровне моря при соответственных колебаниях атмосферного давления и парциального давления кислорода в альвеолярном воздухе гемоглобин практически полностью насыщается кислородом. В условиях высокогорья, где атмосферное давление низкое, снижается парциальное давление кислорода и уменьшается кислородная емкость крови. На содержание кислорода в крови также влияет температура крови: с повышением температуры тела снижается кислородная насыщенность крови. Высокое содержание в крови водородных ионов и диоксида углерода способствует отщеплению кислорода от оксигемогло-бина при прохождении крови через капилляры большого круга кровообращения. Обмен газов между кровью и тканями совершается так же, как и обмен газов между кровью и альвеолярным воздухом — по законам диффузии и осмоса. Поступающая сюда артериальная кровь насыщена кислородом, его напряжение составляет 100 мм рт. ст. В тканевой жидкости напряжение кислорода составляет 20...37 мм рт. ст., а в клетках, которые потребляют кислород, его уровень падает до 0. Поэтому оксигемоглобин отщепляет кислород, который переходит сначала в тканевую жидкость, а затем в клетки тканей.
В процессе тканевого дыхания из клеток выделяется диоксид углерода. Он сначала растворяется в тканевой жидкости и создает там напряжение около 60...70 мм рт. ст., что выше, чем в крови (40 мм рт. ст.). Градиент напряжения кислорода в тканевой жидкости и крови является причиной диффузии диоксида углерода из тканевой жидкости в кровь.
20 — 3389
Поступающий из тканей диоксид углерода незначительно растворяется в плазме крови —до 2,5об.%; его растворимость немного выше, чем у кислорода. Из плазмы диоксид углерода проникает в эритроциты и вытесняет из оксигемоглобина кислород. Оксигемоглобин превращается в восстановленнный, или редуцированный, гемоглобин. Присутствующий в эритроцитах фермент4 карбоангидраза ускоряет соединение диоксида углерода с водой и образование угольной кислоты — Н2С03. Эта кислота нестойкая, она диссоциирует на Н+ и HCOJ. Поскольку мембрана эритроцита непроницаема для Н+, он остается в эритроцитах, а НС03 переходит в плазму крови, где превращается в бикарбонат натрия (NaHC03). Часть диоксида углерода в эритроцитах соединяется с гемоглобином, образуя карбогемоглобин, а с катионами калия — бикарбонат калия (КНС03). В легочных альвеолах, где парциальное давление диоксида углерода ниже, чем в венозной крови, растворенный и освободившийся при диссоциации карбогемоглобина диоксид углерода диффундирует в альвеолярный воздух. Одновременно кислород переходит в кровь и связывается с редуцированным гемоглобином, образуя оксигемоглобин. Оксигемоглобин, являясь более сильной кислотой, чем угольная, вытесняет угольную кислоту из бикарбонатов ионы калия. Угольная кислота расщепляется до С02 и Н20 при участии карбоангидразы. Диоксид углерода переходит из эритроцитов в плазму крови и затем в альвеолярный воздух (см. рис. 7.6).
Несмотря на то что основная часть диоксида углерода присутствует в плазме крови в форме бикарбоната натрия, в альвеолярный воздух выделяется преимущественно диоксид углерода не из плазмы крови, а из эритроцитов. Дело в том, что только в эритроцитах имеется карбоангидраза, расщепляющая угольную кислоту. В плазме крови карбоангидразы нет, поэтому бикарбонаты разрушаются очень медленно и диоксид углерода не успевает выйти в альвеолярный воздух (по легочным капиллярам кровь проходит менее чем за 1 с). Таким образом, диоксид углерода находится в крови в трех формах: растворенной, в виде карбогемоглобина, бикарбонатов, но через легкие удаляется только в одной форме — С02. Не весь кислород из артериальной крови поступает в ткани, часть его переходит в венозную кровь. Отношение объема кислорода, поглощенного тканями, к содержанию его в артериальной крови называется коэффициентом утилизации кислорода. В условиях физиологического покоя он составляет около 40 %. При более высоком уровне метаболизма коэффициент утилизации кислорода увеличивается и уровень его в венозной крови падает. Проходя через легкие, не весь диоксид углерода поступает в альвеолярный воздух, часть его остается в крови и переходит в арте- риальную кровь. Таким образом, если в венозной крови содержится 58 об.% диоксида углерода, то в артериальной крови — 52 об.%. Наличие определенного уровня кислорода и особенно диоксида углерода в артериальной крови имеет огромное значение в процессах регуляции внешнего дыхания. Тканевое (внутриклеточное) дыхание. Тканевое дыхание — это процесс биологического окисления в клетках и тканях организма. Биологическое окисление происходит в митохондриях. Внутреннее пространство митохондрий окружено двумя мембранами—наружной и внутренней. На внутренней мембране, имеющей складчатое строение, сосредоточено большое количество ферментов. Поступающий в клетку кислород затрачивается на окисление жиров, углеводов и белков. При этом освобождается энергия в наиболее доступной для клеток форме, прежде всего в форме АТФ — аденозинтрифосфорной кислоты. Ведущее значение в окислительных процессах имеют реакции дегидрирования (отдача водорода). Синтез АТФ осуществляется при миграции электронов от субстрата к кислороду через цепь дыхательных ферментов (фла-виновые ферменты, цитохромы и др.) Освобождающаяся энергия накапливается в форме макроэргических соединений (например, АТФ), а конечными продуктами реакций становятся вода и диоксид углерода. Наряду с окислительным фосфорилированием кислород может использоваться в некоторых тканях по типу непосредственного внедрения в окисляемое вещество. Такое окисление называется микросомальным, ибо происходит в микросомах — везикулах, образованных мембранами эндоплазматического ретику-лума клетки. Ткани и органы имеют разную потребность в кислороде: интенсивнее поглощают кислород из крови головной мозг, особенно кора больших полушарий, печень, сердце, почки. Меньше потребляют кислорода в состоянии покоя клетки крови, скелетные мышцы, селезенка. При нагрузке потребление кислорода возрастает. Например, при тяжелой мышечной работе скелетные мышцы потребляют больше кислорода в 40 раз, сердечная мышца — в 4 раза (в расчете на 1г ткани).
Даже в пределах одного органа потребление кислорода может резко отличаться. Например, в корковой части почек оно интенсивнее, чем в мозговой части, в 20 раз. Это зависит от строения ткани, плотности распределения в ней кровеносных капилляров, регуляции кровотока, коэффициента утилизации кислорода и ряда других факторов. Следует помнить, что чем больше клетки будут потреблять кислорода, тем больше образуется продуктов обмена — диоксида углерода и воды.
20*
Главная биологическая функция дыхания — это обеспечение газообмена в тканях. Именно ради тканевого дыхания в процессе эволюции возникли и совершенствовались системы кровообращения и внешнего дыхания. Доставка тканям кислорода, выведение водородных ионов и диоксида углерода должны точно соответствовать потребностям тканей и организма в определенный период их жизнедеятельности. В реализации этих процессов, их динамического равновесия участвуют сложные механизмы, включающие регуляцию газового состава крови, регионального кровообращения и трофику тканей. В данной главе мы рассмотрим, каким образом организм поддерживает определенное содержание кислорода и диоксида углерода в крови, т. е. каким образом регулируются дыхательные циклы, глубина и частота дыхания. Внешнее дыхание регулируется нейрогуморальными механизмами. Еще в 1885 г. русский физиолог Н. А. Миславский обнаружил в продолговатом мозге дыхательный центр и доказал наличие в нем двух отделов — центр вдоха и центр выдоха. Центробежными (эфферентными) путями дыхательный центр связан с эффекторами — дыхательными мышцами. Афферентные, или сенсорные, или центростремительные, импульсы поступают в дыхательный центр от различных экстеро- и интерорецепторов, а также от вышележащих отделов головного мозга. Таким образом, в виде довольно упрощенной общей схемы можно представить типичную рефлекторную дугу, состоящую из рецепторов, афферентных путей, нервного центра, эфферентных путей и эффекторов — дыхательных мышц. Дыхательный центр. Дыхательный центр — это совокупность нейронов, расположенных во всех отделах ЦНС и принимающих то или иное участие в регуляции дыхания. Главная часть, или как принято говорить, «ядро» дыхательного центра, находится, как доказал экспериментально Миславский, в продолговатом мозге, в области ретикулярной формации на дне четвертого мозгового желудочка. Без этого отдела дыхание невозможно, повреждение продолговатого мозга приводит неминуемо к смерти из-за остановки дыхания.
Четкий морфологический раздел между центрами вдоха и выдоха в продолговатом мозге отсутствует, но существует распределение функций между нейронами: одни нейроны — инспиратор-ные — генерируют потенциалы действия, возбуждающие инспи-раторные мышцы, другие — экспираторные — возбуждают выдыхательные мышцы. В инспираторных нейронах электрическая активность включается быстро, постепенно нарастает частота им-пульсации (до 70... 100 импульсов в 1 с) и резко падает к концу вдоха. Эта импульсация приводит к сокращению диафрагмы, межреберных и других инспираторных мышц. «Выключение» инспираторных нейронов приводит к расслаблению инспираторных мышц и выдоху. Активность экспираторных нейронов при нормальном спокойном дыхании имеет меньшее значение. Но при усиленном дыхании, особенно при форсированном выдохе, экспираторные нейроны определяют сокращение выдыхательных мышц. Буль барный отдел дыхательного центра головного мозга обладает автоматией. Эта уникальная особенность дыхательного центра заключается в том, что его нейроны могут спонтанно, т. е. самопроизвольно, без каких-либо внешних воздействий деполяризовываться, или разряжаться. Впервые спонтанные колебания электрической активности дыхательного центра обнаружил И. М. Сеченов. Природа автоматии дыхательного центра до сих пор не выяснена. Вероятно, она зависит от специфики обмена веществ нейронов этой области головного мозга и особой чувствительности инспираторных нейронов к окружающей среде, составу цереброспинальной жидкости. Автоматия дыхательного центра сохраняется после почти полной его деаффе-рентации, т. е. после прекращения воздействий со стороны различных рецепторов. Благодаря автоматии дыхательный центр продолговатого мозга обеспечивает ритмичные чередования вдоха и выдоха и определяет частоту дыхания в условиях физиологического покоя. Бульбарный отдел дыхательного центра является самым устойчивым отделом ЦНС к действию наркотических препаратов. Даже при глубоком наркозе, когда рефлекторные реакции отсутствуют, самостоятельное дыхание сохраняется. В арсенале фармакологических препаратов имеются вещества, избирательно повышающие возбудимость дыхательного центра, — лобелии, цититон, воздействующие на дыхательный центр рефлекторно, через рецепторы синокаротидной зоны. И. П. Павлов говорил, что дыхательный центр, который раньше представляли величиной с булавочную головку, необычайно разросся: он спустился вниз в спинной мозг и поднялся вверх до коры больших полушарий. Какую же роль играют другие отделы дыхательного центра? В спинном мозге находятся нейроны (мотонейроны), иннерви-руюшие дыхательные мышцы (рис. 7.7). Возбуждение к ним передается от инспираторных и экспираторных нейронов продолговатого мозга по нисходящим проводящим путям, лежащим в белом веществе спинного мозга. В отличие от бульварного центра мотонейроны спинного мозга не обладают автоматией. Поэтому после перерезки спинного мозга сразу за продолговатым дыхание останавливается, так как дыхательные мышцы не получают команды к сокращениям. Если же спинной мозг перерезать на уровне 4...5-го шейного позвонка, то самостоятельное
Рис. 7.7. Схема организации центрального аппарата регуляции дыхания дыхание сохраняется за счет сокращений диафрагмы, потому что центр диафрагмального нерва расположен в 3...5-M шейных сегментах спинного мозга. Выше продолговатого мозга, прилегая к нему, находится варолиев мост, в котором расположен «пневмотаксический центр». Он не обладает автоматией, но благодаря непрерывной активности обеспечивает периодическую деятельность дыхатель- ного центра, увеличивает скорость развития инспираторной и экспираторной импульсаций в нейронах продолговатого мозга. Средний мозг имеет большое значение в регуляции тонуса поперечнополосатых мышц. Поэтому при сокращении различных мышц афферентная импульсация от них поступает в средний мозг, который соответственно мышечной нагрузке изменяет характер дыхания. Средний мозг ответствен также за координацию дыхания с актами глотания, рвоты и отрыгивания. Во время глотания дыхание задерживается на фазе выдоха, надгортанник закрывает вход в гортань. При рвоте, отрыгивании газов происходит «холостой вдох» — вдох при закрытой гортани. При этом сильно снижается внутриплевральное давление, что и способствует поступлению содержимого из желудка в грудную часть пищевода. Гипоталамус — отдел промежуточного мозга. Значение гипоталамуса в регуляции дыхания заключается в том, что в нем содержатся центры, контролирующие все виды обмена веществ (белковый, жировой, углеводный, минеральный), и центр тепло-регуляции. Поэтому усиление обмена веществ, повышение температуры тела ведут к усилению дыхания. Например, при повышении температуры тела дыхание учащается, что способствует увеличению отдачи теплоты вместе с выдыхаемым воздухом и предохраняет организм от перегревания (тепловая одышка). Гипоталамус принимает участие в изменении характера ды Многогранно значение коры больших полушарий головного мозга в регуляции дыхания. В коре расположены центральные отделы всех анализаторов, информирующих как о внешних воздействиях, так и о состоянии внутренней среды организма. Поэтому наиболее тонкое приспособление дыхания к сиюминутным потребностям организма осуществляется при обязательном участии высших отделов нервной системы. Особое значение имеет кора больших полушарий при мышечной работе. Известно, что учащение дыхания начинается за несколько секунд до начала работы, сразу после команды «приготовиться». Аналогичное явление наблюдается у спортивных лошадей наряду с тахикардией. Причиной подобных «опережающих» реакций у людей и животных являются выработавшиеся в результате повторных тренировок условные рефлексы. Только влиянием коры больших полушарий можно объяснить произвольные, волевые изменения ритма, частоты и глубины дыхания. Человек может произвольно задержать дыхание на несколько секунд или усилить его. Несомненна роль коры в изменении паттерна дыхания во время подачи голоса, при нырянии, обнюхивании. Итак, в регуляции внешнего дыхания участвует дыхательный центр. Ядро этого центра, находящееся в продолговатом мозге, посылает ритмичные импульсы через спинной мозг к дыхательным мышцам. Сам же бульварный отдел дыхательного центра находится под постоянным воздействием со стороны вышележащих отделов ЦНС и различных рецепторов — пульмональных, сосудистых, мышечных и др. Значение рецепторов легких в регуляции дыхания. В легких имеются три группы рецепторов: растяжения и спадения; ирритантные; юкстакапиллярные. Рецепторы растяжения расположены между гладкими мышцами в воздухоносных путях — вокруг трахеи, бронхов и бронхиол, а в альвеолах и плевре отсутствуют. Растяжение легких при вдохе вызывает возбуждение механорецепторов. Возникающие потенциалы действия передаются по центростремительным волокнам блуждающего нерва в продолговатый мозг. К концу вдоха частота импульсации нарастает от 30 до 100 импульсов в 1 с и становится пессимальной, вызывая торможение центра вдоха. Начинается выдох. Рецепторы спадения легких изучены недостаточно. Возможно, при спокойном дыхании их значение невелико. Рефлексы с механорецепторов легких названы по имени открывших их ученых — рефлексы Геринга — Брейера. Назначение этих рефлексов заключается в следующем: информировать дыхательный центр о состоянии легких, их наполненности воздухом и в соответствии с этим регулировать последовательность вдоха и выдоха, ограничивать чрезмерное растяжение легких при вдохе или спадение легких при выдохе. У новорожденных рефлексы с механорецепторов легких играют большую роль; с возрастом значение их уменьшается.
Таким образом, значение блуждающего нерва в регуляции дыхания заключается в передаче афферентных импульсов от механорецепторов легких в дыхательный центр. У животных после перерезки вагуса информация от легких не попадает в продолговатый мозг, поэтому дыхание становится медленным, с коротким вдохом и очень продолжительным выдохом (рис. 7.8). При раздражении вагуса наблюдается задержка дыхания в зависимости от того, в какую фазу дыхательного
цикла действует раздражение. Если раздражение поступает во время вдоха, то вдох преждевременно прекращается и сменяется выдохом, а если совпадает с фазой выдоха, то, наоборот, выдох сменяется вдохом (рис. 7.9). Ирритантные рецепторы расположены в эпителиальном и субэпителиальном слоях всех воздухоносных путей. Они раздражаются при попадании в воздухоносные пути пыли, ядовитых газов, а также при достаточно больших изменениях объема легких. Некоторая часть ирритантных рецепторов возбуждается при обычных вдохах и выдохах. Рефлексы с ирритантных рецепторов носят защитный характер — чихание, кашель, глубокий вдох («вздох»). Центры данных рефлексов расположены в продолговатом мозге. Юкстакапиллярные рецепторы (юкста — вокруг) находятся вблизи капилляров малого круга кровообращения. По функциям они сходны с рецепторами спадения, для них раздражителем является увеличение интерстициального пространства легких, например при отеке. Раздражение юкстакапилляр-ных рецепторов вызывает одышку. Возможно, при интенсивной мышечной работе кровяное давление в легочных сосудах повышается, это увеличивает объем интерстициальной жидкости и стимулирует активность юкстакапиллярных рецепторов. Раздражителем пульмональных рецепторов может быть гистамин, синтезирующийся в базофилах и тучных клетках. В легких этих клеток довольно много, и при аллергических заболеваниях они выделяют гистамин в таком количестве, что это приводит к отеку и одышке. Значение рецепторов дыхательных мышц. В дыхательных мышцах имеются рецепторы растяжения — мышечные веретена, сухожильные рецепторы. Особенно велика плотность их размещения в межреберных мышцах и мышцах стенок живота. Механорецепто-ры дыхательных мышц возбуждаются при их сокращении или растяжении при вдохе или выдохе. По принципу обратной связи они регулируют возбуждение мотонейронов спинного мозга в зависимости от их исходной длины и сопротивления, которое они встре-
Значение хеморецепторов в регуляции дыхания. Исключительно важное значение в регуляции внешнего дыхания имеет газовый состав артериальной крови. Биологическая целесообразность этого вполне понятна, поскольку от содержания кислорода и диоксида углерода в артериальной крови зависит обмен газов между кровью и тканями. Давно стали классикой опыты Фредерика (1890) с перекрестным кровообращением, когда артериальная кровь от одной собаки поступала в кровь другой, а венозная кровь от головы второй собаки — в венозную кровь первой собаки (рис. 7.10). Если пережать трахею и тем самым остановить дыхание первой собаки, то ее кровь с недостаточным содержанием кислорода и избыточным диоксида углерода омывает головной мозг второй собаки. Дыхательный центр второй собаки усиливает дыхание (гипер-пноэ), и в ее крови снижается концентрация диоксида углерода и урежается дыхание вплоть до остановки (апноэ). Благодаря опытам Фредерика стало очевидным, что дыхательный центр чувствителен к уровню содержания газов в артериальной крови. Возросшая концентрация диоксида углерода (гипер-капния) и водородных ионов в крови вызывает учащение дыхания, вследствие чего диоксид углерода выделяется с выдыхаемым воздухом и его концентрация в крови восстанавливается. Снижение содержания диоксида углерода в крови (гипокапния), напротив, вызывает урежение дыхания или его остановку до тех пор, пока в крови концентрация диоксида углерода снова не достигнет нормальной величины (нормокапния). Концентрация кислорода в крови также влияет на возбудимость дыхательного центра, но в меньшей мере, чем диоксида углерода. Это связано с тем, что при обычных колебаниях атмосферного давления, даже на высотах до 2000 м над уровнем моря, почти весь гемоглобин превращается в оксигемоглобин, поэтому парциальное давление кислорода в артериальной крови всегда выше, чем в тканевой жидкости, и ткани получают, во всяком случае в
состоянии физиологического покоя, достаточно кислорода. При значительном снижении парциального давления кислорода в воздухе уменьшается содержание кислорода в крови (гипоксемия) и в тканях (гипоксия), в результате этого возбудимость дыхательного центра повышается и дыхание учащается. Снижение концентрации кис-Рис. 7.10. Перекрестное кровообращение лорода В крови (гипоксемия) МО- жет произойти и вследствие более интенсивного потребления его тканями. В этом случае возможно развитие кислородной недостаточности, что, в свою очередь, вызовет усиление внешнего дыхания. При повышении содержания кислорода в крови, например при вдыхании газовой смеси с высоким содержанием кислорода или при нахождении в барокамере под высоким атмосферном давлении, вентиляция легких уменьшается за счет угнетения дыхательного центра. Мы рассмотрели в отдельности значение содержания кислорода и диоксида углерода в артериальной крови, т. е. аналитически. Однако в действительности оба газа влияют на дыхательный центр одновременно. Установлено, что гипоксия повышает чувствительность дыхательного центра к повышенному содержанию диоксида углерода, и усиление дыхания в этих условиях является интегральной реакцией дыхательного центра в ответ на изменение газового состава крови. Так, при физической работе в мышцы поступает больше кислорода из притекающей крови, увеличивается коэффициент утилизации кислорода, а его концентрация в крови снижается. Одновременно в результате повышения метаболизма из мышц в кровь поступает больше углекислоты и органических кислот. Велика роль сосудистых хеморецепторов при первом вдохе новорожденного. Снижение содержания кислорода в крови и увеличение диоксида углерода во время родов, особенно после пережатия пуповины, является главнейшим раздражителем дыхательного центра, что и вызывает первый вдох. Если в течение 1 мин произвольно максимально усилить дыхание и вызвать этим гипервентиляцию легких, то заметно удлиняется дыхательная пауза между выдохом и последующим вдохом. Может наступить кратковременное апноэ — остановка дыхания на 1...2 мин. Без предшествующей гипервентиляции задержать дыхание можно лишь на 20...30 с. Подобную гипервентиляцию легких с последующим апноэ вызывают у себя ныряльщики — охотники за жемчугом или губкой. После длительных тренировок они остаются под водой до 4...5 мин. Попробуем разобраться в механизмах апноэ после одышки. Поскольку при обычном спокойном дыхании кровь насыщена кислородом на 95 %, усиление дыхания не приводит к значительному увеличению концентрации кислорода в крови. На содержание же диоксида углерода гипервентиляция оказывает заметное влияние — уровень диоксида углерода снижается сначала в альвеолярном воздухе, а затем в крови. Следовательно, апноэ после гипервентиляции легких связано с уменьшением концентрации углекислоты в крови. Дыхание восстановится, когда в крови снова накопится достаточный, или пороговый, уровень диоксида углерода. Если задержать дыхание на 20...30 с, то наступает неудержимое стремление вздохнуть и сделать несколько глубоких дыхательных движений. Следовательно, задержка ведет к гиперпноэ — усиле-
нию дыхания. Это также обусловлено накоплением в крови диоксида углерода, так как за 20...30 с концентрация кислорода в крови снизится незначительно, а диоксид углерода постоянно поступает в кровь из тканей. Итак, диоксид углерода является главнейшим гуморальным раздражителем дыхательного центра. Изменение его концентрации в крови ведет к таким изменениям в частоте и глубине дыхания, которые восстанавливают постоянный уровень углекислоты в крови. При увеличении уровня диоксида углерода в крови происходит стимуляция дыхательного центра и усиление дыхания, при снижении — уменьшение частоты и глубины дыхания. Поэтому столь эффективен метод искусственного дыхания «изо рта в рот», а в газовые смеси для искусственного дыхания обязательно добавляют диоксид углерода. Где же находятся те датчики, или рецепторы, которые улавливают концентрацию газов в крови? Они расположены там, где необходим тщательный контроль за газовым составом внутренней среды организма. Такими участками являются сосудистые рефлексогенные зоны каротидного синуса и аорты, а также центральные рефлексогенные зоны в продолговатом мозге. Синокаротидная зона, или зона каротидного синуса, имеет особо важное значение в отслеживании газового состава и рН крови. Она находится в области разветвления сонных артерий на наружные и внутренние ветви, откуда артериальная кровь направляется в головной мозг. Пороговая концентрация кислорода, углекислоты и водородных ионов для рецепторов синокаротидной зоны соответствует их уровню в крови при нормальных условиях в состоянии покоя. Небольшое возбуждение возникает в отдельных рецепторах при редком глубоком дыхании, когда концентрация газов в крови начинает немного изменяться. Чем сильнее изменяется газовый состав крови, тем большая частота импульсации возникает в хеморецепторах, стимулируя дыхательный центр. Изменение дыхательных движений происходит не только при раздражении хеморецепторов аорты или каротидного синуса. Раздражение находящихся здесь же баро- или прессорецепторов при повышении артериального давления обычно ведет к замедлению дыхания, а при снижении артериального давления — к его усилению. Однако при физической нагрузке повышение артериального давления не приводит к угнетению дыхания, а также к депрессор-ным рефлексам. Центральные (медуллярные) хеморецепторы в продолговатом мозге чувствительны к уровню содержания диоксида углерода в цереброспинальной жидкости. Если артериальные хеморецепторы регулируют газовый состав артериальной крови, то центральные хеморецепторы держат под контролем газовый и кислотно-щелочной гомеостаз жидкости, омывающей головной мозг, — наиболее уязвимую ткань организма. Хеморецепторы, чувствительные к из- менению рН, диоксида углерода и кислорода, имеются также в венозных сосудах и в различных тканях организма. Однако их значение заключается не в регуляции внешнего дыхания, а в изменении регионального, или местного, кровотока. Большой интерес представляют механизмы изменения дыхания при физической работе: при большой нагрузке частота и сила дыхательных движений увеличиваются, что приводит к гипервентиляции легких. Что является причиной этого? Усиление тканевого дыхания в мышцах приводит к накоплению молочной кислоты до Ю0...200мг/100мл крови (вместо 15...24 в норме) и недостатку кислорода для окислительных процессов. Такое состояние называется кислородной задолженностью. Молочная кислота, являясь более сильной кислотой, чем угольная, вытесняет из бикарбонатов крови диоксид углерода, в результате этого возникает гиперкапния, что усиливает возбудимость дыхательного центра. Далее при мышечной работе возбуждаются различные рецепторы: проприорецепторы мышц и сухожилий, механорецепторы легких и воздухоносных путей, хеморецепторы сосудистых рефлексогенных зон, рецепторы сердца и др. От этих и других рецепторов афферентная импульсация также достигает дыхательного центра. При мышечной работе повышается тонус симпатического отдела нервной системы, увеличивается содержание катехоламинов в крови, которые стимулируют дыхательный центр и рефлекторно, и непосредственно. При мышечной работе увеличивается теплопродукция, что также ведет к усилению дыхания (тепловая одышка). Раздражение различных экстерорецепторов приводит к образованию условных рефлексов. Обстановка, в которой обычно совершается работа (ипподром, ландшафт, взнуздывание, появление наездника, а также время суток), является комплексным стереотипом раздражения, подготавливающим лошадь к последующей работе. Наряду с различными поведенческими актами у животного заранее усиливается работа сердца, повышается артериальное давление, перестраивается дыхание и возникают другие вегетативные изменения. В начале работы энергия мышцам поставляется за счет анаэробных процессов. В дальнейшем этого оказывается недостаточно и тогда возникает новое стационарное состояние («второе дыхание»), при котором увеличивается вентиляция легких, систолический и минутный объем сердца, кровоток в работающих мышцах. Таким образом, регуляция дыхания включает два механизма: регуляцию внешнего дыхания, направленную на обеспечение оптимального содержания кислорода и диоксида углерода в крови, т. е. адекватного тканевому метаболизму, и регуляцию кровообращения, создающую наилучшие условия обмена газов между кровью и тканями.
ОСОБЕННОСТИ ДЫХАНИЯ У ПТИЦ Особенности дыхания у птиц обусловлены их образом жизни — полетом и своеобразием анатомического строения дыхательных путей. Эти особенности свойственны всему классу птиц — как летающих, так и нелетающих. У птиц сильно развита грудная клетка, большая грудная кость, вместо реберных хрящей грудные костные ребра подвижно соединены с позвоночными ребрами. Диафрагма у птиц редуцирована, поэтому не имеет большого значения в дыхании. Наружная поверхность легких вдавлена между ребрами и прочно срастается с ними. Свободная поверхность легких гладкая, покрыта плеврой.
Трахея входит в легкие, разветвляется на бронхи и бронхиолы. В бронхиолах происходит обмен газов с притекающей сюда венозной кровью. Часть бронхов выходит из легких и заканчивается воздухоносными мешками —это тонкостенные образования, заполненные воздухом (рис. 7.11). Они расположены между органами, соединяются с бронхами, а некоторые из них тонкими трубочками соединяются с во
|
||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-16; просмотров: 843; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.112.91 (0.02 с.) |

 Транспорт диоксида углерода кровью. Диоксид углерода транспортируется в трех формах: в растворенном виде, в соединении с гемоглобином (карбогемоглобин) и в виде бикарбонатов.
Транспорт диоксида углерода кровью. Диоксид углерода транспортируется в трех формах: в растворенном виде, в соединении с гемоглобином (карбогемоглобин) и в виде бикарбонатов. 7.4. РЕГУЛЯЦИЯ ДЫХАНИЯ
7.4. РЕГУЛЯЦИЯ ДЫХАНИЯ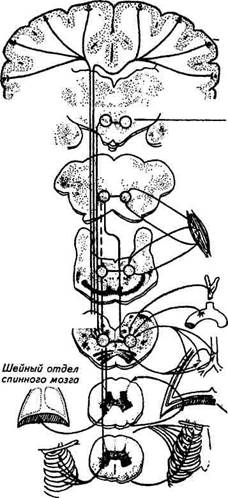

 чают при сокращении. Сильное раздражение механорецепторов грудной клетки (например, при ее сжатии) вызывает торможение инспираторной деятельности дыхательного центра.
чают при сокращении. Сильное раздражение механорецепторов грудной клетки (например, при ее сжатии) вызывает торможение инспираторной деятельности дыхательного центра.
 В регуляции вдоха и выдоха большее значение имеют авто-матия дыхательного центра и афферентные импульсы от меха-норецепторов легких и дыхательных мышц, а в регуляции частоты и глубины дыхания — газовый состав крови, цереброспинальной жидкости и афферентные импульсы от хеморецепто-ров кровеносных сосудов, тканей и медуллярных (бульбарных) хеморецепторов.
В регуляции вдоха и выдоха большее значение имеют авто-матия дыхательного центра и афферентные импульсы от меха-норецепторов легких и дыхательных мышц, а в регуляции частоты и глубины дыхания — газовый состав крови, цереброспинальной жидкости и афферентные импульсы от хеморецепто-ров кровеносных сосудов, тканей и медуллярных (бульбарных) хеморецепторов.



