
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Пищеварение в тонком кишечникеСодержание книги
Поиск на нашем сайте В результате пищеварения в тонком кишечнике происходит расщепление сложных питательных веществ до тех остатков (в основном мономеров), в которых они могут всосаться в кровь и лимфу. В кишечнике осуществляется окончательный гидролиз пищевых веществ под действием поджелудочного и кишечного соков, желчи и всасывание продуктов гидролиза. К тонкому отделу кишечника относят двенадцатиперстную, тонкую и подвздошную кишки. В полость двенадцатиперстной кишки открываются протоки поджелудочной железы и желчный проток. Натощак содержимое двенадцатиперстной кишки имеет слабощелочную реакцию (рН 7,2...8,0). При поступлении в нее порций желудочного содержимого реакция среды становится кислой, однако быстро происходит ее нейтрализация за счет желчи, поджелудочного сока и кишечного сока, которые ингибируют желудочный пепсин. Чем выше кислотность дуоденального содержимого, тем больше выделяется поджелудочного сока и желчи. Поджелудочный сок. Роль поджелудочной железы в пищеварительном процессе определяется ее внешнесекреторной деятельностью, т. е. выделением поджелудочного сока в просвет двенадцатиперстной кишки. Количество выделяемого за сутки поджелудочного сока в среднем составляет у жвачных 6...7 л, у свиней 8 л, у собаки 200...300 мл, у человека около 1 л. Поджелудочный сок представляет собой бесцветную жидкость щелочной реакции, обусловленной высокой концентрацией анионов HCOJ. Кроме того, в нем присутствуют хлорид натрия, хлорид кальция, фосфаты и другие неорганические ионы.
Трипсин — основной протеолитический фермент поджелудочного сока. Первоначально выделяется в виде неактивного профермента трипсиногена, который активируется ферментом, вырабатываемым слизистой оболочкой двенадцатиперстной кишки, — энтеропептидазой (энтерокиназой). Трипсин активен в щелочной среде, гидролизует белки и их промежуточные соединения — альбумозы и пептоны — до полипептидов, дипептидов и даже аминокислот. Химотрипсин выделяется также в неактивном состоянии в виде химотрипсиногена и активируется трипсином; расщепляет белки преимущественно после их обработки пепсином и трипсином. Панкреатопептидаза (эластаза) осуществляет гидролиз специфических белков соединительной ткани и му-кополисахаридов, расщепляя их на пептиды и аминокислоты. Карбоксипептидаза отщепляет от пептидов свободные аминокислоты со стороны карбоксильной группы. Дезокси-рибонуклеаза и рибонуклеаза осуществляют гидролиз нуклеиновых кислот. Липаза — липолитический фермент поджелудочного сока. Обладает широким диапазоном действия на жиры, расщепляя их на глицерин и жирные кислоты. Растворяется в воде и действует на жиры только на границе раздела вода — жир. Липаза активируется ионами кальция и желчными кислотами. К гликолитическим (или амилолитическим) ферментам поджелудочного сока относятся: амилаза — расщепляет крахмал, гликоген и амилопектин на декстрины и мальтозу; глюкози-даза (мальтаза) — расщепляет мальтозу на две молекулы глюкозы; фруктофуронидаза — расщепляет сахарозу на глюкозу и фруктозу. У молодняка в молочный период в поджелудочном соке отмечается высокое содержание галактозидазы, расщепляющей молочный сахар лактозу на глюкозу и галактозу. Секреция поджелудочной железы. Регуляция осуществляется с участием двух основных механизмов — нервно-рефлекторного и гуморального. Естественным стимулятором секреции служит прием пищи. Первая фаза секреции поджелудочной железы — сложно-рефлекторная — обусловлена одновременным действием условных (вид и запах пищи) и безусловных (жевание и глотание пищи) раздражителей. Афферентные пути от этих рецепторов аналогичны афферентным путям слюноотделительного рефлекса и рефлекса желудочной секреции. Эфферентные пути рефлексов поджелудочной секреции от центра, расположенного в продолговатом мозге, проходят в составе блуждающих нервов. Раздражение блуждающего нерва оказывает слабое стимулирующее влияние на секрецию поджелудочной железы, однако при этом секрет железы существенно обогащается ферментами. Секреция поджелудочного сока во время гуморальной фазы определяется гормонами желудочно-кишечного тракта, прежде всего секретином и холецистокинином (или панкреозимином). Секретин образуется в неактивном состоянии — в виде про-секретина. В полости двенадцатиперстной кишки он активируется соляной кислотой и другими кислыми веществами, затем всасывается в кровь и оказывает стимулирующее действие на клетки поджелудочной железы. Секрецию поджелудочного сока, кроме того, стимулируют гастрин, серотонин, инсулин, вазоактивный интес-тинальный пептид, соли желчных кислот. Холецистоки-н и н приводит к увеличению количества ферментов в поджелудочном соке при сохранении его объема. Тормозящим эффектом обладают глюкагон, простагландины, желудочный тормозной полипептид, кальцитонин. Желчеобразование и желчевьщеление. Секреция желчи осуществляется клетками печеночной паренхимы, собранными в печеночные дольки. От печеночных клеток начинаются желчные капилляры, которые объединяются в желчные протоки. Последние, сливаясь между собой, образуют печеночный проток, открывающийся в желчный пузырь. У лошади, верблюда, оленя желчный пузырь отсутствует, а его функцию выполняют хорошо развитые желчные ходы. Общее количество выделяющейся желчи за сутки составляет у лошади и жвачных около 6 л, у овец — 300...400 мл, у собаки — 250...300 мл. В желчеобразовании различают три процесса: активная секреция, например билирубина, желчных кислот; фильтрация из плазмы крови воды и растворенных в ней веществ: глюкозы, креатинина, ионов калия, натрия, хлора; реабсорбция, т. е. всасывание обратно в кровь некоторых элементов первичной или печеночной желчи. Таким образом, часть веществ, входящих в состав желчи, синтезируется клетками печени, другие поступают в нее из плазмы крови. В желчном пузыре происходит реабсорбция из желчи некоторых солей и воды, в результате чего из печеночной желчи образуется более густая, концентрированная, так называемая пузырная желчь. Плотность печеночной желчи составляет 1,009...1,013, содержание плотных веществ 1...5 %, рН 7,5. Пузырная желчь содержит 9...20% плотных веществ; плотность ее 1,026...1,048, рН 6,8. Кроме того, в ее состав входит слизь, выделяющаяся бокаловидными клетками слизистой оболочки желчного пузыря. Желчь в печени образуется непрерывно, однако сфинктер желчного протока находится в сокращенном состоянии до тех пор, пока в полость кишечника не поступит пищевая кашица.
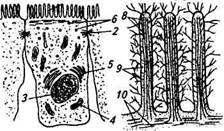
Желчь, поступающая в кишечник, способствует пищеварению за счет своей щелочной реакции путем нейтрализации кислого химуса, поступающего из желудка. Натриевые соли желчных кислот уменьшают поверхностное натяжение воды и жира, способствуя эмульгированию жиров пищевой кашицы, при этом их поверхность увеличивается и облегчается процесс взаимодействия с липолитическими ферментами. Кроме того, соли желчных кислот активируют липазу поджелудочного сока и ускоряют всасывание конечных продуктов расщепления жира. Желчные кислоты усиливают моторику кишечника; соединяясь с нерастворимыми в воде жирными кислотами, образуют водорастворимые комплексы, всасывающиеся в кишечнике. Желчеобразование в клетках печени происходит непрерывно, однако оно может быть стимулировано актом еды, раздражением блуждающего нерва или гуморальным путем (например, действие секретина). Поступление желчи в двенадцатиперстную кишку регулируется нервно-рефлекторным и гуморальными механизмами. Первые порции желчи поступают в кишечник через 5... 10 мин после приема пиши. Для накопления и выделения желчи важна синхронная работа мышц сфинктера желчного протока и мышц желчного пузыря. При эвакуации желчи сокращение стенок пузыря сопровождается расслаблением сфинктера. При накоплении желчи наблюдается обратный процесс — расслабление мышц стенок пузыря и сокращение сфинктера. Эта согласованная работа мышц регулируется нервно-рефлекторным и гуморальным путями. Раздражение блуждающего нерва приводит к сокращению мышц пузыря и расслаблению круговой мышцы сфинктера, т. е. эвакуации желчи. Стимуляция симпатического нерва прекращает поступление желчи в кишечник. Гуморальная регуляция выделения желчи связана с выработкой в слизистой оболочке двенадцатиперстной кишки гормона холе-цистокинина под влиянием промежуточных продуктов гидролиза белков и жиров. Холецистокинин вызывает сокращение мышц пузыря и расслабление мышцы сфинктера желчного протока. Важный гуморальный возбудитель желчевыделения — и сама желчь: ее поступление в кишечник способствует желчеобразованию. Структурно-функциональная организация эпителиального слоя тонкого кишечника. Для эпителия, выстилающего полость кишечника, характерна складчатая поверхность, проявляющаяся на разных уровнях организации кишечными складками, ворсинками и микроворсинками апикальной мембраны эпителиальных клеток (рис. 5.9). Кишечные ворсинки увеличивают поверхность эпителия в 8 раз, а микроворсинки — в 30...60 раз.
Рис. 5.9. Схема строения слизистой обо- Рис. 5.10. Схема строения кишечной Лочки тонкой кишки ворсинки Основной структурной единицей слизистой тонкой кишки является ворсинка, имеющая свой сосудистый, мышечный и нервный аппарат (рис. 5.10). Каждая ворсинка пронизана густой сетью кровеносных капилляров, в ней также находятся несколько лимфатических синусов. Капилляры ворсинок тесно контактируют с эпителиальными клетками, что облегчает путь пищеварительных субстратов и обмен между эпителием и кровеносной системой. В строме ворсинок находятся гладкомышечные клетки, обеспечивающие их продольное сокращение. Подобные ритмические сокращения ворсинок способствуют микроциркуляции и продвижению питательных веществ. Поверхность кишечных ворсинок покрывает однослойный цилиндрический эпителий с ярко выраженной полярностью. Клетки эпителиального пласта представлены на 90 % энтероцитами с характерной исчерченной (или щеточной) каемкой, образованной микроворсинками апикальной плазматической мембраны (рис. 5.11). Кроме них в составе кишечного пласта встречаются бокаловидные клетки, секретирующие мукополисахариды, а также эндокринные клетки и клетки Панета, содержащие ацидофильные гранулы. В составе ^-1. М,^7>^ Рис. 5.11. Схема строения всасывающей клетки (А) кишечного эпителия и ее щеточной каемки (Б): 1 — микроворсинки; 2 — десмосома; 3 — ядро; 4— митохондрии; 5— аппарат Голь-джи; 6— эндоцитозные везикулы; 7— гли-кокаликс; 8— актиновые филаменты; 9— филаменты а-актинина; 10— нити миозина
Рис. 5.12. Схема регуляции пищеварительных процессов на уровне эпителиального пласта:
эпителиального пласта присутствуют лимфоциты, проникающие в строму ворсинок через базальную мембрану, количество которых существенно варьирует в зависимости от физиологического состояния желудочно-кишечного тракта. Слизистую оболочку тонкой кишки покрывает непрерывный слой так называемых слизистых наложений. В его состав входят слизь, фрагменты слущенных эпителиоцитов, мембранные везикулы, отдельные лейкоциты. Здесь могут также находиться пищевые частицы, бактерии, простейшие. От физико-химических свойств слизи зависит скорость проникновения через слой слизистых наложений тех или иных веществ. Доказано содержание в слизистом слое ряда гидролитических ферментов и его сорб-ционные свойства. Таким образом, слой слизистых наложений выполняет защитную (механическую и химическую), буферную, пищеварительную и транспортную функции. Эпителиальный пласт кишечника — чрезвычайно сложная саморегулирующаяся система, находящаяся под строгим нервно-гуморальным контролем. Простейший механизм регуляции его функций локализован в пределах самого эпителия (рис. 5.12). Роль рецепторов играют нейроэндокринные клетки (совмещающие функции нервных и эндокринных клеток), которые в случае изменения тех или иных условий в полости кишечника приходят в состояние возбуждения и выделяют в базальную часть эпителиального пласта биологически активные вещества. Эти биологически активные вещества (например, серотонин) могут действовать непосредственно на клетки эпителия, изменяя их всасывающую, секреторную и другие функции. Кроме того, в регуляции функций кишечного эпителия могут участвовать клетки интрамуральных нервных ганглиев (рис. 5.13). Регистрируемое раздражение (изменение параметров внутрики-шечного содержимого) либо по нервно-синаптическим связям, либо дистантно посредством биологически активных веществ вызывает активацию нейронов подслизистого и межмышечного сплетений. Выделяемые нейронами классические медиаторы и вещества типа коротких пептидов действуют на клетки эпителиального слоя, приводя к изменению его функционального состояния. Рис. 5.13. Схема регуляции пищеваритель- Внешнее
ментов подслизистого сплетения: 1 — эпителиоцит; 2— нейроэндокринная клетка; 3 — бокаловидная клетка; 4— афферентный нейрон; 5— вставочный нейрон; 6— моторный нейрон в составе подслизистого сплетения Состав и значение кишечного сока. В слизистой оболочке тонкого кишечника содержатся ли-беркюновы железы, вырабатывающие кишечный пищеварительный сок. Кишечный сок —бесцветная жидкость, которая при отстаивании разделяется на два слоя: нижний, содержащий слизистые комочки, и верхний — жидкий прозрачный. Слизистые комочки состоят из секрета бокаловидных желез и слущенных эпителиальных клеток, на которых адсорбировано до 70...80 % ферментов. Кишечный сок обладает протеолитической, липоли-тической и амилолитической активностью. Энтеропептидаза (энтерокиназа) продуцируется в начальной части тонкого отдела кишечника. Она гидролизует трипсиноген и прокарбоксипептйдазу, превращая их в активные ферменты. Действие ее на другие белки ограничено вследствие высокой специфичности. Аминопептидаза, аминотрипептидаза и другие кишечные пептидазы расщепляют в основном пептиды, образующиеся в результате действия пепсина и трипсина. Пептидазы расщепляют пептиды до свободных аминокислот. Кишечный сок гидролизует нативные белки, за исключением казеина. Щелочная фосфатаза принимает участие в отщеплении фосфатидов от различных соединений и фосфорилирова-нии углеводов, аминокислот и липидов, обеспечивая их транспорт через клеточные мембраны. Щелочная фосфатаза присутствует почти во всех тканях организма, но в эпителиальных клетках ворсинок тонкого кишечника ее в 30...40 раз больше, чем в печени или поджелудочной железе. В кишечном соке имеются все ферменты, действующие на углеводы. Но особенно высока активность ферментов, расщепляющих дисахариды: глюкозидазы, фруктофуронидазы,
По сравнению с ферментами желудочного или поджелудочного сока ферменты кишечного сока действуют на продукты промежуточного гидролиза питательных веществ. Так, пептидазы кишечного сока не действуют на нативные белки или высокомолекулярные продукты их разложения, но разлагают пептиды невысокой молекулярной массы до отдельных аминокислот. Основным возбуждающим фактором в регуляции образования и выделения кишечного сока является сама пищевая кашица — химус. Вероятно, нервно-рефлекторная регуляция выделения кишечного сока осуществляется за счет нервных сплетений (Мейс-нерова и Ауэрбахова) в стенке кишечника. Симпатическая и парасимпатическая регуляции осуществляются посредством чревного и блуждающего нервов. Гуморальная регуляция сокоотделения в тонком кишечнике контролируется возбуждающими (вазоактив-ный кишечный полипептид, энтерокинин, холецистокинин, гаст-рин) и тормозящими (желудочный тормозной полипептид, секретин) гормонами. Пристеночное (мембранное) пищеварение. В тонком кишечнике происходят заключительные этапы расщепления питательных веществ и всасывание конечных продуктов гидролиза. Ведущее место в осуществлении этих процессов занимает пристеночное, или мембранное, пищеварение, осуществляемое ферментными системами, локализованными на границе вне- и внутриклеточной среды. Необходимое для этого увеличение площади поверхности, на которой могут идти пищеварительные процессы, создается благодаря наличию специфических выростов плазматической мембраны энтероцитов — микроворсинок, которые образуют так называемую щеточную каемку. На поверхности микроворсинок, в свою очередь, имеется слой гликопротеидов — гликокаликс (см. рис. 5.11). Таким образом, достичь мембраны, т. е. места локализации пищеварительных ферментов, могут только небольшого размера молекулы. Поэтому пристеночное пищеварение наиболее эффективно для промежуточных продуктов гидролиза. Кроме того, оно происходит в стерильных условиях, поскольку бактерии и другие микроорганизмы не способны проникнуть через сеть гликокаликса. Это одно из основных отличий пристеночного пищеварения от полостного. Другая особенность — это место локализации (топография). В отличие от полостного пищеварения, наиболее эффективного в двенадцатиперстной кишке, эффект пристеночного пищеварения максимален в тонкой кишке. Пристеночное пищеварение сопряжено с процессами всасывания питательных веществ. Питательные вещества всасываются в основном в виде мономеров. Причем если они представляют собой продукты расщепления олигомеров, то всасывание их происходит быстрее, чем просто мономеров. Например, глюкоза, введенная в кишку, всасывается медленнее, чем глюкоза, которая образуется как продукт гидролиза введенного в кишку крахмала. Это говорит о сопряженности на мембранах энтероцитов процессов гидролиза и транспорта и активизации транспорта процессами гидролиза. Ферменты пристеночного пищеварения имеют двоякое происхождение: частично они адсорбируются из химуса, а частично синтезируются в самих эпителиальных клетках. На поверхности мембраны ферменты и соответственно их активные центры локализованы в определенном порядке, что повышает эффективность процессов гидролиза. В щеточной кайме обнаружены щелочная фосфатаза, аминопептидаза, липаза, амилаза, фруктофуронидаза и другие ферменты. Моторика тонкого кишечника. Моторная деятельность кишечника обеспечивает перемешивание пристеночного слоя химуса, повышает внутриклеточное давление, усиливает всасывание пищеварительных веществ и передвижение содержимого по пищеварительному тракту. Моторика кишечника осуществляется благодаря координированным сокращениям и расслаблениям наружного продольного и внутреннего циркулярного слоев мышц. Волокна гладких мышц кишечника обладают автома-тией, т. е. свойством ритмически сокращаться без воздействия внешних раздражителей. Известно четыре типа кишечных сокращений: ритмическая сегментация, перистальтика, маятникообразные и тонические сокращения (рис. 5.14).
Характер моторики перистальтика
ритмическая сегментация маятникообразные движения
тоническое сокращение Рис. 5.14. Виды моторики кишечника Ритмической сегментацией называются ритмические (8... 10 раз в минуту) сокращения слоя циркулярных мышц, в результате чего возникают перетяжки (шириной 1...2 см) и участки расслабления мышц между этими перетяжками (шириной 15...20 см). Перетяжки сегментируют кишку, делят ее содержимое на части. Растяжение расслабленного участка возбуждает миоци-ты, что сопровождается образованием новой перетяжки. При этом расслабляются мышцы ранее образованной перетяжки. Перистальтические сокращения подобны волне, распространяющейся вдоль по кишке и продвигающей ее содержимое. Волна начинается с сокращения циркулярных мышц выше порции химуса и растяжения продольных мышц ниже этой порции. Вдоль кишки могут одновременно идти несколько волн перистальтических сокращений. Слабые волны перемешивают поверхностный слой химуса вблизи стенки кишки. Мощные перистальтические волны могут распространяться по тонкой кишке вплоть до илеоцекальной заслонки и довольно быстро освобождать кишечник от пищевых масс. Маятникообразные движения возникают при сокращении продольного, в меньшей степени циркулярного мышечного слоя. Такие движения перемещают химус «вперед-назад». Чередование ритмической сегментации и маятникообразных движений способствует тщательному перемешиванию пищевых масс. Тонические сокращения обусловлены определенным тонусом гладких мышц тонкой кишки. Они составляют основу перистальтического, маятникообразного сокращений и ритмической сегментации. Отсутствие тонуса мышц кишечника (атония) исключает возможность любого вида сокращений. Ворсинки кишечника также постоянно сокращаются и расслабляются, что способствует обновлению химуса, всасыванию и оттоку лимфы. Двигательная активность тонкого кишечника регулируется нервно-рефлекторным и гуморальным механизмами. Сокращение мышц вызывается механическим и химическим раздражением слизистой оболочки пищевой кашицей. Гуморальная регуляция двигательной активности тонкого кишечника связана с действием ацетилхолина, энтерокинина и серотонина. Кроме того, моторику возбуждают экстрактивные вещества, желчь, соли кальция и магния.
|
|||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-16; просмотров: 471; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.011 с.) |


 В поджелудочном соке содержатся ферменты, действующие на все группы питательных веществ: протеолитические, амило-литические и липолитические.
В поджелудочном соке содержатся ферменты, действующие на все группы питательных веществ: протеолитические, амило-литические и липолитические.
 В состав желчи входят вода, слизь и пигменты (билирубин и биливердин), желчные кислоты (холевая, дезоксихолевая, литохо-левая), желчнокислые соли, холестерин и лецитин, соли неорганических кислот. Ферменты в желчи отсутствуют.
В состав желчи входят вода, слизь и пигменты (билирубин и биливердин), желчные кислоты (холевая, дезоксихолевая, литохо-левая), желчнокислые соли, холестерин и лецитин, соли неорганических кислот. Ферменты в желчи отсутствуют.
 1 — эпителиоциты; 2 — нейроэндокринная клетка; 3 — бокаловидная секреторная клетка
1 — эпителиоциты; 2 — нейроэндокринная клетка; 3 — бокаловидная секреторная клетка пых процессов с участием нервных эле- воздействие
пых процессов с участием нервных эле- воздействие галактозидазы. Кишечная липаза расщепляет жиры, но ее содержание в кишечном соке незначительно. Ф о с -фолипаза действует на эфирные связи в фосфолипидах, расщепляя их на жирные кислоты, глицерин и фосфаты.
галактозидазы. Кишечная липаза расщепляет жиры, но ее содержание в кишечном соке незначительно. Ф о с -фолипаза действует на эфирные связи в фосфолипидах, расщепляя их на жирные кислоты, глицерин и фосфаты.


