
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механизмы взаимодействия гормона с клеткамиСодержание книги
Поиск на нашем сайте
Гормоны способны оказывать ряд физиологических действий. Метаболическое, связанное с изменением обмена веществ. Большинство гормонов участвуют в регуляции обмена веществ путем изменения активности ферментативных систем в тканях. Для некоторых гормонов воздействие на обменные процессы является основной функцией. Например, инсулин, глкжа-гон и адреналин непосредственно регулируют углеводный обмен; глюкокортикоиды стимулируют образование углеводов из продуктов распада белков, минералкортикоиды влияют на содержание натрия и калия в организме, а гормон околощитовидной железы регулирует обмен кальция и фосфора. Соматотропный гормон гипофиза стимулирует синтез белка и расходование углеводов и жиров. Влияние на уровень энергетических процессов гормонов щитовидной железы реализуется за счет усиления расщепления белков, углеводов и жиров. Влияние половых гормонов проявляется в усилении синтеза белков (особенно мышечных) и интенсивном расходовании жира и изменении минерального обмена. Морфогенетическое, связанное с изменением дифференциации клеток и тканей, ростом и метаморфозом. Хорошо известно действие гормонов щитовидной железы на метаморфоз головастиков; нарушение ее функций приводит к ощутимым нарушениям роста и развития молодняка большинства животных. Гона-дотропные гормоны гипофиза стимулируют рост, развитие и диф-ференцировку клеток половых желез. Начало продукции гормонов половыми железами обусловливает развитие половых органов и появление вторичных половых признаков. Кинетическое, или пусковое, воздействие, вызывающее Коррегирующее, связанное с изменением уровня интенсивности функций организма или его органов, работающих и без участия гормонального влияния. При этом происходит усиление или ослабление физиологических процессов. Так, адреналин учащает ритм и увеличивает силу сердечных сокращений, но угнетает сократительную активность мускулатуры желудочно-кишечного тракта.
Ключевой этап в реализации физиологического действия гормона на клетку-мишень — это его связывание со специфическим белком-рецептором, который служит распознающим посредником гормонального эффекта. Наличие рецептора — необходимое условие развития эндокринной функции, и, если в клетке нет рецепторов, гормон не способен воздействовать на нее. В общем виде рецептор для любого из гормонов состоит из трех пространственно обособленных структур: осуществляющей избирательный прием гормонального сигнала за счет специфического и обратимого связывания гормона; осуществляющей преобразование внешнего гормонального сигнала во внутриклеточный сигнал; ответственной за инициацию регуляторных эффектов гормона за счет взаимодействия гормонорецепторного комплекса с различными акцепторными участками клетки (рис. 12.1). Таким образом, рецептор — это такая химическая структура соответствующей ткани-мишени, которая имеет высокоспецифичные участки для связывания гормональных соединений, причем в результате этого связывания инициируются последующие биохимические реакции, необходимые для осуществления конечного эффекта данного гормона.
Рис. 12.1. Модель функционально-структурной организации рецепторной молекулы: а — гормоносвязывающий участок; е — эффекторный участок; зигзагообразная линия — участок сопряжения а и е; А — акцептор; Г— гормон
ными акцепторными структурами. Как правило, мембранным акцептором служит либо аденилатциклаза, изменяющая уровень внутриклеточного циклического аденозинмонофосфата (цАМФ), либо неэлектрогенные кальциевые каналы, изменяющие внутриклеточную концентрацию ионизированного кальция, либо специфические протеазы, способные изменить внутриклеточное содержание специфических гликопептидов. Существует и возможность внутриклеточного действия связанного с мембраной гормона. Мембранные рецепторы обычно асимметрично встроены в плазматическую мембрану, причем связывающий гормонузнающий фрагмент их молекул жестко ориентирован к наружной поверхности и обращен к внеклеточному пространству. Взаимодействующая с акцептором часть ре-цепторной молекулы (или рецепторного комплекса) ориентирована в противоположном направлении и обращена к цитоплазме клетки. Эта жесткая ориентация рецепторов обеспечивает век-торность трансмембранной передачи гормональной информации внутрь клеток. Локализуясь трансмембранно, гидрофобная часть мембранных (поверхностных) рецепторов оказывается погруженной в бислойную липидную мембрану, в то время как связывающие и исполнительные участки находятся в водной фазе. Несмотря на относительную жесткость фиксации таких рецепторов в мембране, они способны перемещаться, осуществляя продольные и поперечные движения, облегчая тем самым «поиск» рецептором гормона. Возможная подвижность рецеп-торных молекул позволяет осуществить клетке такие процессы, как агрегирование и интернализация рецепторов, связавших гормональные молекулы. В основе этого процесса лежит специальный процесс — адсорбтивный эндоцитоз. Диффузно распределенные по клеточной поверхности рецепторы после связывания гормона приобретают способность к движениям в латеральной плоскости. Сближаясь, они объединяются (кластеризуются) в особых участках плазматической мембраны — окаймленных ямках — местах начала интернализации. По данным электронной микроскопии, окаймленные ямки представляют собой углубления в клеточной мембране со стороны цитоплазмы, выстланные слоем щетинкоподобного белка — клатрина, выполняющего функцию своеобразного каркаса (клатриновая корзинка). Загруженные кластерами гормонорецепторных комплексов окаймленные ямки постепенно углубляются, замыкаются и отрываются от мембраны, превращаясь в свободно плавающие по цито-плазматическому пространству везикулы. В ходе перемещения по цитоплазме происходит утрата клатринового покрытия, гладкие везикулы объединяются, формируя рецептосомы, которые способны сливаться с лизосомами или структурами аппарата Гольджи. Образование комплексной эндолизосомальной везикулы приводит к ферментативному разделению комплекса гормон-рецептор,
Рис. 12.2. Схема рецепторопосредован- ^ — ё > ё ного эндоцитоза: ( /—окаймленная ямка; //— эндосома; 111— | мультивезикулярные эндосомы; АГ— аппарат I
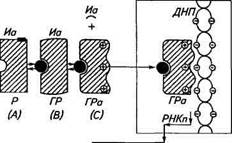
причем «проникший» в составе эндосомы гормон способен связываться с внутриклеточными структурами, а рецептор имеет возможность вновь встроиться в плазматическую мембрану (ре-циклировать) (рис. 12.2). Рецепция стероидных гормонов. Стероидные гормоны как липо-фильные вещества способны сравнительно свободно проникать через плазматические мембраны внутрь клетки и быстро связываться с соответствующим цитозольным рецептором. Образованный в цитоплазме гормонорецепторный комплекс способен активироваться за счет освобождения от внутриклеточных ингибиторов. Процесс активации сопровождается дополнительной кон-формационной перестройкой молекулы рецептора с локальным выходом на его поверхность положительных зарядов — остатков лизина и аргинина, что позволяет ему электростатически связываться с полианионами, к которым, в частности, относится ДНК. Электростатическое и структурное взаимодействие гормон-рецеп-торного комплекса с хроматином, происходящее в результате транслокации его в ядро, приводит к индукции множественных эффектов гормона, регулируя процессы транскрипции. При этом возможно сосуществование двух механизмов:
селективного — легко насыщаемое избирательное связывание комплексов с ограниченным числом специфических акцепторных участков ДНК, ответственных за транскрипцию определенных генов; определяющего интенсивность процесса — ненасыщаемое связывание комплексов с большим числом разных участков ДНК и других компонентов хроматина, приводящих к их де-конденсации и общему повышению матричной активности. Гормонорецепторный комплекс, взаимодействуя с хроматином, специфически изменяет уровень синтеза определенных мРНК, рРНК и выход их за пределы ядра, а также синтез структурных и функциональных белков (рис. 12.3). На заключительном этапе необходимо присутствие факторов терминации, инактивиру-ющих рецепторы, и ферментов стероидного метаболизма, разрушающих гормон, что позволяет завершить рабочий цикл рецепторов.
Г Г
Гормональные эффекты Рис. 12.3. Модель основных стадий рецепции стероидных гормонов: Г— гормон, Р— рецептор; ГР— исходный гормонорецепторный комплекс; ГРа — активированный гормонорецепторный комплекс; А, В, С— конформационные состояния гормонорецепторно-го комплекса; ДНП— хроматин; РНКп — индуцируемые РНК Рецепция тиреоидных гормонов. Освобожденные от транспортных плазматических белков гормоны щитовидной железы — три-йодтиронин и тироксин — относительно свободно проникают внутрь клеток. В цитоплазме тироксин, превращаясь в трийодти-ронин, связывается со специфическими ядерными рецепторами, которые располагаются на молекуле ДНК. Связывание гормон-рецепторного комплекса зависит как от ионного окружения, так и от особенностей первичной и вторичной структуры акцепторной ДНК. Функциональное сопряжение гормонорецепторного комплекса с ДНК зависит от гистонов, способных повысить сродство рецепторов к гормону и сродство комплекса к специфическим локусам ДНК, что и обеспечивает избирательную регуляцию синтеза соответствующих функциональных мРНК и рРНК. В ходе трансляции индуцированные РНК меняют уровень синтеза кодируемых ими белков.
Рецепция белково-пептидных гормонов. В отличие от стероидных и тиреоидных гормонов белково-пептидные гормоны и кате-холамины действуют на гормонозависимые клетки через поверхностные рецепторы, локализованные в плазматических мембранах. Обнаружено три самостоятельных пути прохождения информационного гормонального сигнала с поверхности клетки: аденилатциклазный; кальциевый; протеазный. Эти пути могут реализоваться самостоятельно, однако в реальных физиологических условиях они тесно сопряжены. Аденилатциклазный механизм реализуется посредством взаимодействия гормонорецепторного комплекса с локализованным в мембране ферментом аденилатциклазой, образующим внутриклеточный посредник цАМФ. Деятельность аденилатциклазы осуществляется при участии факторов трансмембранного сопряжения: гуанозинтрифосфат (ГТФ) — связывающий N-белок, локализованный в липидном слое мембраны; ГТФ, катионы магния и марганца и анионы железа, компоненты цитоскелета (микротрубочки и микрофиламенты); кислые липи-ды мембран и некоторые ферменты составляют аденилатциклаз-ную систему. ГТФ, играющий основную роль в трансмембранном сопряжении, связывает N-белок, и этот комплекс активирует аде-нилатциклазу, которая способна при образовании одного гормон-рецепторного комплекса образовать до 500 молекул цАМФ. В комплексе с рецептором и аденилатциклазой N-белок способен присоединять к себе ГТФазу, которая, расщепляя ГТФ, инактивируя N-белок и аденилатциклазу, прекращает процесс активации цик-лазного фермента. Внутриклеточная концентрация посредника 3', 5'-мононуклеотида (цАМФ) регулируется цитоплазматическим ферментом фосфодиэстеразой, расщепляющей его до неактивного 5'-мононуклеотида (рис. 12.4).
Белки хроматина
Необходимо отметить, что стимулирующие эффекты гормонов могут вторично усиливаться и распространяться за счет образования простагландинов — производных арахидоновой кислоты, которая под влиянием циклооксигеназы превращается в циклические эндоперекиси PgG2 и PgH2, из которых затем образуются про-стагландины. Простагландины, выходя из материнской клетки, могут действовать на те же или соседние клетки и также стимулировать аденилатциклазную систему.
|
||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-16; просмотров: 361; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.1.23 (0.014 с.) |

 Исходя из локализации рецепторов, характера акцепторных участков и особенностей гормонзависимых эффектов, рецепцию разделяют на внутриклеточную и мембранную. Внутриклеточная рецепция характерна для стероидных и тиреоидных гормонов. Гормон свободно проникает через плазматическую мембрану внутрь клетки и взаимодействует с цитозольными, ядерными и другими рецепторами. Мембраносвязанные рецепторы связывают гормоны на поверхности клеток, и гормональные эффекты развиваются благодаря образованию внутриклеточных посредников (медиаторов) в результате взаимодействия гормоноре-цепторных комплексов с мембран-
Исходя из локализации рецепторов, характера акцепторных участков и особенностей гормонзависимых эффектов, рецепцию разделяют на внутриклеточную и мембранную. Внутриклеточная рецепция характерна для стероидных и тиреоидных гормонов. Гормон свободно проникает через плазматическую мембрану внутрь клетки и взаимодействует с цитозольными, ядерными и другими рецепторами. Мембраносвязанные рецепторы связывают гормоны на поверхности клеток, и гормональные эффекты развиваются благодаря образованию внутриклеточных посредников (медиаторов) в результате взаимодействия гормоноре-цепторных комплексов с мембран-

 Цитозоль
Цитозоль Биологический эффект
Биологический эффект Образовавшийся в клетке цАМФ специфически взаимодействует с регуляторными субъединицами цАМФ-зависимых протеин-киназ, происходит диссоциация молекул фермента на мономеры. В результате каталитическая субъединица активизируется и обеспечивает в цитоплазме фосфорилирование целого ряда синтезированных функциональных и структурных белков, изменяя их функциональное состояние: активируются киназа фосфорилазы, липаза, фосфолипаза, фосфопротеины рибосом, гистоны и т. д. Кроме того, комплекс цАМФ с одной из регуляторных субъединиц транслоцируется в клеточное ядро и оказывает избирательное воздействие на активность генома в ходе транскрипции.
Образовавшийся в клетке цАМФ специфически взаимодействует с регуляторными субъединицами цАМФ-зависимых протеин-киназ, происходит диссоциация молекул фермента на мономеры. В результате каталитическая субъединица активизируется и обеспечивает в цитоплазме фосфорилирование целого ряда синтезированных функциональных и структурных белков, изменяя их функциональное состояние: активируются киназа фосфорилазы, липаза, фосфолипаза, фосфопротеины рибосом, гистоны и т. д. Кроме того, комплекс цАМФ с одной из регуляторных субъединиц транслоцируется в клеточное ядро и оказывает избирательное воздействие на активность генома в ходе транскрипции.


