
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Сухожильный рецептор ГольджиСодержание книги
Поиск на нашем сайте Другие важнейшие афферентные рецепторы двигательного анализатора—сухожильные рецепторы Гольджи (см. рис. 15.2), расположенные главным образом на границе мышечной и сухожильной тканей. Сухожильные рецепторы напоминают по форме мышечные веретена. Функцию вспомогательного аппарата выполняют сухожильные волокна, покрытые соединительнотканной оболочкой — капсулой. Сухожильные волокна внутри капсулы лежат не так плотно, как снаружи, и имеют меньший диаметр. К рецептору подходит одиночное миелинизированное волокно диаметром 8...15мкм, которое внутри капсулы многократно делится, теряя миелиновую оболочку; его тонкие кустиковидные окончания располагаются на сухожильных волокнах. Эфферентных окончаний в отличие от мышечных веретен сухожильные рецепторы Гольджи не имеют. Электрическая реакция рецепторов Гольджи на растяжение следующая: рецепторный потенциал и последовательность потенциалов действия напоминает таковую вторичных афферентных окончаний мышечных веретен (см. рис. 15.3), т. е. сухожильные рецепторы Гольджи являются медленноадаптирующимися рецепторами растяжения — детекторами амплитуды. РЕФЛЕКС НА РАСТЯЖЕНИЕ МЫШЦЫ Рассмотрим некоторые простые примеры функционирования двигательного анализатора с участием мышечных веретен и рецепторов Гольджи. В формировании миотатического рефлекса, или рефлекса на растяжение мышц (рис. 15.5), принимают участие афферентные нейроны, образующие в мышечных веретенах пер-
В г Рис.15.5. Рефлекс на растяжение мышцы: А. В исходном «заданном» состоянии груз небольшой массы (1) удерживается экстра-фузальными волокнами мышцы. В нервных волокнах, образующих афферентные окончания, регистрируются только редкие спонтанные потенциалы действия. Б. При увеличении массы груза (2 > 1) мышца с мышечными веретенами растягивается. В афферентных волокнах повышается частота потенциалов действия, которые поступают через синап-тические контакты на а-мотонейроны (показано стрелкой в направлении от мышечного веретена) и возбуждают их. От а-мотонейронов потенциалы действия направляются к экс-трафузальным мышечным волокнам (стрелки в направлении к мышце) и через синаптичес-кие контакты вызывают сокращение мышцы. В. Сокращение мышцы произошло не до заданной длины. Ликвидация «ошибки» осуществляется с помощью фузимоторных гамма-нейронов, образующих двигательные окончания на интрафузальных мышечных волокнах веретен. Г. Мышца возвращается к заданной длине 41—3389 вичные афферентные окончания, и ос-мотонейроны, которые обеспечивают двигательную иннервацию экстрафузальных мышечных волокон. При растяжении мышцы происходит растяжение и мышечных веретен, что сопровождается повышением частоты потенциалов действия в афферентных волокнах. Поскольку афферентные нейроны синаптически связаны в ЦНС с а-мотонейронами, то в последних также повышается частота потенциалов действия. Распространяясь по эфферентным волокнам, потенциалы действия через синаптические окончания вызывают сокращение — укорочение длины мышцы. Уменьшение растягивающего воздействия на интрафузальные волокна снижает частоту потенциалов действия в афферентных нервных волокнах, и система возвращается в состояние, близкое к исходному. Однако полного восстановления первоначальной длины данная система не обеспечивает. Оставшуюся небольшую разницу между исходной длиной мышцы (до растяжения) и длиной после рефлекторного сокращения (ее называют ошибкой) система не может определить. Для этого потребовалось бы звено обратной связи, т. е. мотонейрон с неограниченно высокой чувствительностью. Способствует возврату мышц к исходной «заданной» длине так называемая фузимоторная система, включающая в себя интрафузальные мышечные волокна и фузимоторные (у) мотонейроны, которые образуют на интрафузальных мышечных волокнах двигательные синапсы. Активация этой системы потенциалами действия из двигательных центров анализатора вызывает сокращение концевых участков веретена и тем самым растяжение центрального несокращаюшегося участка, где находятся афферентные первичные окончания. Это приведет к дополнительному увеличению частоты потенциалов действия в афферентном нейроне, которое будет восприниматься а-мотонейро-ном с последующей посылкой к синаптическим окончаниям экстрафузальных волокон эфферентных потенциалов действия. В результате этого в мышце произойдет дополнительное сокращение и исходная длина будет достигнута. Из вышеизложенного становится ясно, что миотатический рефлекс служит для поддержания постоянной длины мышцы при изменениях воздействующей на нее нагрузки. Этот механизм у животных, как, по-видимому, и у человека, осуществляется без контроля сознания и играет определяющую роль в поддержании позы. Ответственные за положение тела в пространстве мышцы-разгибатели должны иметь определенную заданную длину и в противовес силы тяжести удерживать конечности животного в выпрямленном состоянии. Сухожильные рецепторы Гольджи соединены с мышечными волокнами последовательно (а не параллельно, как мышечные веретена), поэтому они должны реагировать на изменение напряжения мышцы, а не длины. Было обнаружено, что через тормозные интернейроны афферентная импульсация i рсигишрмп I <>iii.*i.*m оказывает влияние на а-мотонейроны, снижая vi>">'< m, us,п> ши ности. Это, например, может проявляться в понижпни... ими потенциалов действия, посылаемых к синапсам >kci|),h|>v ини.мы* мышечных волокон, что предотвращает чрезмерное пшфнжгнж мышцы. Предполагается также, что сигнализация сухожильными рецепторами о мышечном напряжении а-мотонейронам cnoioit ствует исправлению неточностей в регуляции длины мышц мин татическими рефлексами. КОЖНЫЙ АНАЛИЗАТОР Кожный анализатор имеет самую большую рецептирующую поверхность, которая превышает в несколько тысяч раз, например, площадь сетчатки глаза, обонятельной области и кортиевого органа слухового анализатора. Кожные ощущения у животных весьма многообразны. К ним в первую очередь можно отнести ощущения легкого прикосновения, давления, вибрации, зуда и щекотки, тепла и холода, боли. Действие каждого раздражения, в свою очередь, имеет множество оттенков. Кожа у сельскохозяйственных животных, за исключением небольших участков, покрыта волосами. Она состоит из двух главных слоев: эпидермиса и дермы. В эпидермисе непрерывно происходят деление клеток и миграция их к поверхности. В ходе этой миграции клетки образуют на поверхности слой мертвых уплощенных клеток — роговой слой, который богат кератином и белком, устойчивым к действию воды и различных химических веществ. Под эпидермисом лежит дерма, представляющая собой мягкую и гибкую структуру благодаря толстому слою из коллагеновых и эластических волокон. Многочисленные и разнообразные рецепторные структуры располагаются в обоих слоях кожи. Кроме того, каждый волос кожи имеет свою афферентную иннервацию. Все рецепторы можно разделить на четыре группы. Для первой группы рецепторов — механорецепторы — адекватным раздражителем является энергия механического стимула, механическое смещение кожи или волос; для второй группы — тепловые рецепторы — повышение температуры кожи; для третьей группы— холодовые рецепторы — понижение температуры. Четвертая группа рецепторов—болевые рецепторы — воспринимает весьма сильные механические и температурные раздражители, вызывающие ощущение боли. Все эти рецепторы образованы разветвлениями нервного волокна (дендрита) первого афферентного нейрона. У одних рецепторов разветвленная часть заключена в одну или несколько капсул (инкапсулированные рецепторы), у других терминали лежат свободно в кожных слоях, но они всегда покрыты шваннов-скими клетками.
Все известные механорецепторы по своим функциональным характеристикам можно разделить на несколько групп. В основе классификации механорецепторов лежит их реакция на сту-пенчатообразные механические стимулы, у которых изменяется амплитуда, скорость ее нарастания и спада, длительность и частота Кроме ступенчатообразных стимулов широко применяются синусоидальные механические стимулы. Механорецепторы входящие в состав какой-либо группы, настроены на восприятие определенного параметра механического стимула.
В коже обнаружены механорецепторы, по своим функциональным характеристикам сходные с мышечными веретенами, т. е. детекторы амплитуды - скорости (рис. 15.6, А). В покое эти рецептор обладаютСпонтанной активностью. В ответ на постоянное
в Рис. 15.6. Электрические ответы кожных механорецепторов на механические стимулы различной формы: А Импульсная активность медленноадаптируюшегося рецептора (дет™ амплитуды-скорости) на ступенчатообразныи механический стимул длительностью около 10 с: /-начало; 2-продолжение, 3- окончание электрического ответа рецептора; калибровка по вертикали 05 мВ, по горизонтали 250 мс. Б. Импульсная активность быст-роадагтшруюшихсяГ механорецепторов (детекторов скорости) на сту-КтообрТзный (I) и пилообразные (2 3, 4) механические стимулы с различной скоростью нарастания; калибровка по вертикали 0,1 мВ, по даонтали для 1 - 125 мс, для 2, 3, 4-250 мс. В Импульсная актив-шсГб^оадаптир^^^^ скоростей) на ступенчатообразный (Г) и пилообразные (2, ^механические стимулы с различной скоростью нарастания; калибровка по вертикали 0,1 мВ, по горизонтали для /, 2— 250 мс, J— n мс смещение кожи (растяжение или надавлииашн-) они inu-pn руют потенциалы действия, частота которых о. ир.-мпи-м уменьшается (медленная адаптация). При увеличении.imihhiv ды раздражающего стимула частота афферентных иокчппын.ш действия повышается и зависимость между силой рачдрилг ния и частотой импульсации носит логарифмический харак тер Импульсная активность, приходящаяся на передний фрош нарастания раздражения, увеличивается с увеличением скорое ти смещения кожи. Параллельные морфологические исследования позволили идентифицировать некоторые из этих рецепторов. Ими оказались инкапсулированные механорецепторы - тельца Руффини и тельца Меркеля, а также большая группа свободных нервных окончаний различной степени ветвления (рис. 15.7). Тельца Руффини располагаются в дерме и представляют собой веретенообразные структуры, окруженные капсулой (см. рис. 15.7, Л). Сложнопе-реплетенные коллагеновые волокна являются вспомогательным аппаратом рецептора, они тесно связаны с коллагеновыми пучками лежащими рядом с тельцем. Миелинизированное волокно диаметром 5...10мкм проникает внутрь капсулы, теряет миели-новую оболочку и многократно делится, образуя кустиковидные окончания, располагающиеся на коллагеновых волокнах. У телец Руффини наиболее выраженная электрическая реакция на растяжение кожи. Тельца Меркеля активней реагируют на тактильное (надавливание на кожу) раздражение. Эти структуры встречаются в волосистой коже и представляют собой куполообразные образования диаметром Ю0...500мкм, возвышающиеся над поверхностью кожи (см. рис. 15.7, Б). Рецепторные окончания локализуются непосредственно под базальной мембраной утолщенного эпидермиса. Миелинизированное волокно (средний диаметр 9 5 мкм) также многократно делится и образует дископодобно расширяющиеся окончания с тесно примыкающей к нему клеткой Меркеля. Нервное окончание содержит большое количество митохондрий. ^^ Кроме медленноадаптирующихся механорецепторов в коже были обнаружены быстроадаптирующиеся рецепторы, которые, как правило, не обладают спонтанной активностью. На постоянное смещение кожи они не генерируют потенциалы действия, какой бы величины оно не было (см. рис. 15.6, Б). Генерация потенциалов действия происходит только во время изменения амплитуды нарастания или спада стимула. С увеличением скорости изменения амплитуды раздражения число потенциалов действия, а следовательно, и их средняя частота увеличиваются. Эти механорецепторы были классифицированы как детекторы скорости. Быстроадаптирующиеся ответы были зарегистрированы от нервных волокон, иннервирующих волосяные луковицы различных типов волос: пуховых, стержневых и вибрисс. Нервные оконча-
Рис. 15.7. Кожные механорецепторы (графическая реконструкция по серийным срезам): А. Кожный рецептор растяжения — тельце Руффини: 1 — афферентное нервное волокно; 2, 5— афферентные нервные окончания; 3 — внутрикапсулярное пространство; 4, 6— колла-геновые волокна; 7—место разветвления афферентного нервного волокна; 8— капсула; 9— шванновская клетка с ядром; калибровка 50 мкм. Б. Тактильное тельце — тельце Меркеля: 1 — тактильная клетка (клетка Меркеля) и примыкающее к ней дископодобное нервное окончание; 2 — эпидермис тактильного тельца; 3 — афферентное нервное волокно; 4— немиели-низированное нервное волокно; калибровка 10 мкм. В. Тельце Пачини: 1— внутренняя часть капсулы с наиболее плотным расположением уплощенных слоев клеток (внутренняя колба); 2— наружняя часть капсулы с менее плотным расположением слоев уплощенных эпителиальных клеток; 3 — афферентное нервное окончание; 4— афферентное нервное волокно; калибровка 600 мкм. Г. Свободные немиелинизированные нервные окончания (7 окончаний), образованные разветвлением миелинизированного нервного волокна и располагающиеся около пухового волоса кожи: 1 — одно из афферентных нервных окончаний (продольный срез); 2— семь нервных окончаний, покрытых шванновской клеткой (поперечный срез); 3 — шванновская клетка; 4— четыре нервных окончания, покрытые шванновской клеткой; 5, 6— терминальные части нервных окончаний, окруженные складками из шванновских клеток (поперечный срез): нервные окончания заполнены мелкими митохондриями; калибровка 1 мкм ния, находящиеся в волосяных фолликулах пуховых и стержневых волос, представляют собой сеть тонких немиелинизиро-ванных окончаний, плотно оплетающих структуры волосяной сумки. Рецепторы волосяной сумки образованы разветвленными (см. рис. 15.7, Г) миелинизированными нервными волокнами диаметром около 6 мкм. Каждый волос иннервируется ответвлениями нескольких волокон. В свою очередь, одно и то же волок- но может иннервировать много волос. Так, например, в ухе кролика одно волокно может давать афферентные веточки примерно к 300 волосяным фолликулам на площади около 1 см2. Вибриссы являются специальными осязательными волосами и обычно длинней и толще, чем другие волосы. У лошадей и коров вибриссы — это длинные торчащие волосы на морде; у кошек, собак и пушных зверей их называют усами. У лазающих животных они находятся на животе. Корень вибриссы окружен кровеносным синусом. Вибриссы сильнее иннервированы, чем обычные волосы. Свыше 100 нервных волокон могут проникать в сумку вибриссы и образовывать внутри афферентные окончания. Кроме быстроадаптиру-ющихся механорецепторов у вибрисс имеются и медленноадапти-рующиеся механорецепторы. Детекторы скорости были обнаружены в различных участках кожи животных, в том числе в коже репродуктивных органов самцов и сосков молочных желез. Морфологически они представляют собой неинкапсулированные нервные окончания различной формы и сложности ветвления. Какова же природа адаптации в этих механорецепторах? Надо отметить,что от детекторов скорости не удалось зарегистрировать рецепторный потенциал, поэтому о причине адаптации нельзя сказать что-либо определенное. Некоторые исследователи счита-ют,что адаптация обусловлена особыми механическими свойствами вспомогательного аппарата рецепторов, который не пропускает статическую компоненту механического стимула. Так обстоит дело с быстроадаптирующимся механорецептором — тельцем Пачини. Этот рецептор реагирует на довольно быстрые изменения скорости смещения кожи всего лишь 1...2 потенциалами действия. Данный тип рецепторов получил название детекторов быстрых скоростей, или детекторов толчков. Тельце Пачини состоит (см. рис. 15.7, В) из нервного окончания, мембрана которого содержит механоактивируемые ионные каналы, и окружающей его капсулы в виде концентрических слоев из уплощенных клеток эпителия, подобных слоям чешуек в луковице. Тельце имеет весьма крупные размеры и может достигать 0,5...0,7 мм в диаметре и 1 мм в длину. Когда тельце при надавливании на него ступенчатообразным механическим стимулом деформируется, эта деформация через капсулу механически передается к мембране нервного окончания. Последняя обычно реагирует кратковременной деполяризацией (рецепторный потенциал), приходящейся на начало и конец раздражения (рис. 15.8, А). Если снять максимально возможное количество оболочек (полностью удалить оболочки капсулы никому не удавалось), то рецепторный потенциал будет приближаться к ступенчатообразной форме (см. рис. 15.8, Б). Была предложена механическая модель капсулы, представляющая ее слои как параллельно и последовательно соединенные вязкоэластичные элементы. Анализ показал, что давление статической (постоянной) части стимула ослабляется
А. Интактное тельце Пачини. На начало и конец раздражения генерируется кратковременный деполяризационный рецепторный потенциал. Б. У тельца Пачини механически удалены внешние слои капсулы. На механический стимул генерируется более длительный рецепторный потенциал примерно в 100 раз при достижении области афферентного окончания. Только быстрый компонент, т. е. изменение скорости раздражения, может проходить через капсулу. Таким образом, капсула играет роль механического фильтра. Надо отметить, что в коже коров, коз, лошадей и других животных было обнаружено большое число как телец Пачини, так и других подобных им рецепторов с различными размерами и количествами слоев капсулы. ТЕРМОРЕЦЕПТОРЫ КОЖИ Способность гомойотермных животных поддерживать постоянную температуру тела несмотря на резкие колебания температуры окружающей среды — одно из важнейших биологических регу-ляторных явлений. Процессы терморегуляции протекают независимо от сознания и не прекращаются во время сна. Начальным звеном терморегулирующей системы кожи являются многочисленные терморецепторы. Поскольку изменения температуры окружающей среды создают субъективные ощущения тепла или холода, то можно предположить, что в коже имеются соответственно тепловые и холодовые рецепторы. Действительно, в первых же объективных исследованиях терморецепции кожи, проведенных в начале 20-х годов XX в., были обнаружены особые точки, в которых при нагревании или охлаждении происходило увеличение частоты афферентных потенциалов действия в нервных волокнах, иннервирующих эти участки. По реакции на адекватный стимул температурные рецепторы были похожи на медленноадаптирую-щиеся механорецепторы кожи (рис. 15.9). В покое у обоих типов терморецепторов была зарегистрирована спонтанная импульсная активность. В ответ на ступенчатообразное изменение температуры (повышение или понижение) частота афферентных потенциалов действия быстро повышалась, достигая максимального значения, и затем сравнительно быстро понижалась до определенного уровня, а далее процесс замедлялся. Действие температурных стимулов с различными скоростями нарастания позволяет отнести
Рис.15.9. Терморецепторы кожи: A. Схематическое изображение одиночного холодового рецептора на безволосой коже носа B. Зависимость между средней частотой импульсной активности холодового (квадраты) и теп температурные рецепторы к детекторам амплитуды скорости. Исследование зависимости частоты афферентных потенциалов действия от величины температуры показало, что средняя частота импульсной активности при нагревании или охлаждении возрастает
Каковы же морфологические корреляты тепловых и Холодовых рецепторов? С конца XIX в. по 60-е годы XX в. существовало мнение, основанное на субъективных данных исследований на человеке, что холодовым рецепторам соответствуют инкапсулированные тельца Краузе, напоминающие по своему строению тельца Пачини, а тепловым — тельца Руффини. Однако объективные исследования с использованием регистрации афферентной импульсации показали, что эти рецепторы являются кожными механо-рецепторами (см. предыдущий раздел). Эксперименты с одновременной регистрацией импульсной активности и морфологической маркировкой выявили, что терморецепторам соответствуют не-инкапсулированные нервные окончания, локализованные в обоих слоях кожи. Например, холодовый рецептор безволосой части носа кошки (см. рис. 15.9, А) представляет собой неразветвлен-ное немиелинизированное окончание в эпидермисе диаметром 2,5...4мкм; расширенный его кончик заполнен мелкими митохондриями. Терморецепторы образованы тонкими миелинизиро-ванными нервными волокнами диаметром 1,5...5 мкм. Изменению частоты афферентных потенциалов действия в нервных волокнах терморецепторов должно предшествовать изменение мембранного потенциала афферентной области (рецепторно-го потенциала) (см. рис. 15.1). Известно, что формирование мембранного потенциала обусловлено пассивным и активным ионным транспортом. Величина пассивной компоненты мембранного потенциала вычисляется с помощью уравнения Голдмана. Поэтому можно предположить, что вариации в температуре будут непосредственно изменять пассивный транспорт ионов и соответственно влиять на амплитуду мембранного потенциала. Однако простые расчеты показывают, что колебание температуры на 10 °С изменяет мембранный потенциал всего лишь на 2 мВ. Если учесть, что терморецепторы обладают спонтанной активностью, т. е. мембранный потенциал находится в области порога генерации потенциалов действия, то данной величины изменения мембранного потенциала все же будет недостаточно для заметного увеличения частоты импульсации. Нужно отметить,что у некоторых видов животных тепловые рецепторы имеют чрезвычайно высокую чувствительность. Так, в лицевой ямке гремучей змеи находятся разветвленные нервные окончания, реагирующие на повышение температуры всего на 0,002 °С. Исследования последних лет позволяют полагать, что в рецепторной мембране имеются каналы, которые активируются повышением или понижением температуры. Дополнительно, что касается рецепции холода, то обширный экспериментальный материал указывает на участие в этом процессе активного ионного транспорта — ионного насоса. В частности, предполагается, что в Холодовых рецепторах при обычной температуре тела животного мембранный потенциал и рецепторных окончаниях сдвинут в сторону гиперпшшрт.шии электрогенным ионным насосом. Работа ионной помпы minimi от температуры, поэтому ее снижение, охлаждение нервной кт> минали будет затормаживать работу помпы, что вызовет деполяризацию мембраны и соответственно увеличение частоты афферентных потенциалов действия. Совокупность тепловых и Холодовых рецепторов обеспечивает функционирование системы терморегуляции организма животного. Так, интенсивное охлаждение тела возбуждает холодо-вые рецепторы. Посредством автономной нервной системы они вызывают сужение кожных кровеносных сосудов, что уменьшает интенсивность кровотока в коже и сохраняет теплоту внутри организма. Снижение теплоотдачи достигается также уменьшением поверхности тела (свертывание в клубок) и изменением положения волос или перьев на теле животного. Одновременно через систему двигательных нервов увеличивается выделение теплоты в мышцах за счет напряжения мышц и мышечной дрожи, а также посредством «разогревающих» движений. Теплопродукция возрастает, кроме того, благодаря усилению обмена веществ во внутренних органах. При внешнем перегревании тела животного тепловые рецепторы кожи вызывают процессы, обратные происходящим при переохлаждении: расширение кровеносных сосудов, усиление потоотделения, учащение дыхания. БОЛЕВЫЕ РЕЦЕПТОРЫ КОЖИ У животных и человека сильные механические или другие воздействия вызывают ощущение боли. Установлено, что в коже сверхинтенсивные стимулы воспринимают специальные болевые рецепторы, которые, как и терморецепторы, образованы тонкими миелинизированными и немиелинизированными нервными волокнами диаметром 0,3...5 мкм. Чрезвычайно важное значение для выяснения природы болевой рецепции имели опыты, проведенные в 80-х годах XX в. на добровольцах с одновременной регистрацией субъективных (речевых) и объективных (афферентных потенциалов действия в нервных волокнах) ответов при раздражении болевыми стимулами. В нормальных условиях в нервных волокнах болевых рецепторов отсутствует спонтанная активность, т. е. в отличие от обычных механо- и терморецепторов мембранный потенциал в зоне генерации потенциалов действия в болевых рецепторах, вероятно, сдвинут от порогового значения в сторону гиперполяризации и для возбуждения необходимо приложить стимул большей амплитуды. Кожные болевые рецепторы делятся
на механоболевые и термоболевые. Есть предположение, что в восприятии болевых стимулов принимают участие химические вещества, выделяющиеся при действие интенсивной стимуляции, — гистамин, брадикинин, а также ионы калия, концентрация которых может повышаться около нервных терминалей при повреждении окружающих их клеточных структур. ОБОНЯТЕЛЬНЫЙ АНАЛИЗАТОР У животных восприятие различных химических веществ, находящихся в окружающей среде, осуществляется с помощью двух химических анализаторов: обонятельного и вкусового. Обонятельный анализатор является дистантным анализатором, т. е. воспринимает химические вещества, источник которых находится на расстоянии (иногда довольно значительном) от животного. Обонятельный анализатор выполняет функцию сигнальной хемо-рецепции. Помимо поиска и выбора пищи, спасения от врагов обоняние помогает при ориентации как на суше, так и в воде. Хорошим примером этого служит возвращение лососевых рыб в родительские водоемы: на заключительных этапах миграции, при попадании лососевых в реки, основным ориентиром для них служит знакомый запах, источник которого находится выше по течению. Существенную роль в жизни животного играет так называемое химическое общение. Оно осуществляется посредством особых химических веществ — «феромонов», которые вырабатываются и выделяются специальными железами. К феромонам относятся половые привлекающие вещества, используемые для ме-чения своей территории. Многие животные по запаху отыскивают или распознают полового партнера. Известно, что запуск барана в стадо овец непосредственно перед сезоном спаривания ускоряет и синхронизирует течку у них. Причина этого заключается в том, что запах барана вызывает у овец более частые выбросы лютеинизирующего гормона, что стимулирует ускоренное развитие фолликулов и овуляцию. Сходное явление наблюдается и у свиней, у которых половое созревание значительно ускоряется при раннем запуске в стадо хряка. Запах детеныша может существенно влиять на проявление материнского инстинкта, что позволяет заключить о существовании обонятельного имприн-тинга как у матери, так и у детеныша. Животных с хорошо развитым обонянием называют макросматшами — это почти все млекопитающие. Значительно слабее развито обоняние у человека и у птиц — это микросматики. Периферическая часть обонятельного анализатора находится в носовой полости: в области верхнего носового хода и в задней верхней части носовой перегородки. Обонятельная область окрашена в более темный цвет: от желто-бурого до темно-корич- Рис. 15.10. Ультраструктурная (А) и функциональная организация (Б) обонятельного рецептора: 1 — обонятельная клетка; 2 —дендрит обонятельной клетки; 3— булавовидное расширение дендрита; 4 — обонятельные волоски (показаны поперечные срезы волосков на различном удалении от булавовидного расширения); 5—опорная клетка; 6—зона плотного контакта; 7— базальная клетка; 8— базальная мембрана; 9— афферентные нервные волокна рецепторных клеток; /—область взаимодействия пахучего вещества с рецепторными белками мембраны волосков и булавовидного расширения дендрита, где происходит активация внутриклеточных посредников (цАМФ, цГМФ, инозитолтрифосфата, ионов кальция); Я—область активации ионных каналов внутриклеточными посредниками и генерация рецепторного потенциала; III— область трансформации рецепторного потенциала в серию потенциалов действия невого в отличие от остальной части носовой полости и представляет собой эпителиальный слой толщиной 150...200 мкм, включающий в себя обонятельные, опорные и базальные клетки (рис. 15.10, Л). Обонятельные клетки являются биполярными нейронами с диаметром сомы 5... 15 мкм (см. рис. 15.10, Б). От сомы клетки к наружной поверхности отходит дендрит диаметром 1...2мкм и длиной 5... 120 мкм. Вершина дендрита образует булавовидное расширение диаметром 2...3 мкм и выступает в слой слизи, покрывающей обонятельную область. От обонятельной «булавы» отходят несколько тонких волосков — цилий, диаметр которых около Ю0...200нм, а длина 5...250 мкм. Отростки обонятельных клеток, идущих в ЦНС (аксоны), принадлежат к группе самых тонких нервных волокон, их диаметр составляет 0,2...0,3 мкм. Число обонятельных клеток у макросматиков может достигать сотен миллионов. Опорные клетки тесно примыкают к обонятельным клеткам и образуют в верхней части зоны плотный контакт (рис. 15.10, А), в результате чего слой слизи отделяется от внеклеточной жидкости, окружающей боковые и базальные участки клеток. На верхней (апикальной) поверхности опорных клеток есть микроволоски, но значительно короче, чем у обонятельных клеток. Базальные клетки лежат в глубине эпителия (см. рис. 15.10, А) и составляют клеточный резерв, из которого образуются рецепторные и опорные клетки. Слизистый слой выступает в качестве вспомогательной структуры у обонятельных рецепторов и имеет большое значение для восприятия химических веществ. В частности, слизь защищает обонятельный эпителий от высыхания у наземных животных, а у водных — от излишнего смачивания. Прежде чем произойдет воздействие молекул пахучего вещества на обонятельные клетки, они должны раствориться в слизи. Нерастворимые вещества не пахнут. Со слизью удаляются остатки пахучих молекул. Источником слизи являются боуменовые железы, находящиеся под эпителиальным слоем. Предполагается, что слизь могут секретировать и опорные клетки. При нормальном дыхании животного через обонятельную область проходит 5... 10 % всего воздуха, остальная же часть поступает в дыхательную систему через нижний носовой ход. Слабые запахи при таком дыхании не воспринимаются. Для того чтобы пахнущие вещества достигли обонятельных рецепторов, необходимо более глубокое дыхание или учащенное. Именно таким образом животное принюхивается, увеличивая ток воздуха через обонятельную полость. Во время еды обонятельные рецепторы раздражаются воздухом, проходящим через хоаны из ротовой полости. Малые размеры обонятельных клеток существенно затрудняют изучение механизмов обоняния на клеточном уровне, много неясного и в вопросе об адекватных раздражителях для обонятельных клеток различных животнных. В окружающем животных воздухе находится множество химических соединений, которые для одних животных биологически значимы и вызывают ощущения, а для других нейтральны и в данной концентрации не вызывают возбуждение рецепторов и не создают в ЦНС никаких ощущений. Поэтому при анализе работы обонятельных клеток необходимо учитывать данный фактор. Благодаря комплексным биохимическим, биофизическим и электрофизиологическим исследованиям выяснили процесс восприятия химического стимула в обонятельных рецепторах (см. рис. 15.1). В отсутствие стимуляции обонятельные клетки генерируют спонтанные афферентные потенциалы действия, что указывает на де-поляризованность мембраны клеток. Измерения показали, что мембранный потенциал обонятельных клеток на 30...50 мВ меньше по абсолютной величине, чем равновесный калиевый потенциал (100 мВ). Таким образом, клеточная мембрана обонятельных клеток в покое проницаема и для других ионов (см. гл. 2). Это зна- чительно увеличивает чувствительность рецептора к действию раздражающего стимула. При вдыхании пахучего вещества обонятельные рецепторы отвечают изменением частоты разрядов афферентных потенциалов действия — увеличением или уменьшением, в зависимости от качества и интенсивности химического стимула, но изменение ее невелико. Например, для наиболее исследованных обонятельных клеток лягушки она редко превышает 10 имп/с. Изменению частоты потенциалов действия в аксонах обонятельных клеток предшествует изменение мембранного потенциала ре-цепторной клетки — рецепторного потенциала. В одних случаях это деполяризация, при которой соответственно происходит увеличение частоты афферентных потенциалов действия в аксоне, в других — мембрана гиперполяризуется, что сопровождается уменьшением частоты спонтанных потенциалов действия или их исчезновением на время действия стимула. Длительность рецепторного потенциала, а следовательно, и длительность последовательности потенциалов действия во много раз превышает длительность химического стимула. По всей видимости, это обусловлено постепенным удалением молекул пахучего вещества из обонятельной полости. Разрушение волосков значительно снижает реакцию обонятельных клеток на химические стимулы. Это позволило сделать заключение
|
|||||
|
|
Последнее изменение этой страницы: 2016-12-16; просмотров: 541; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.021 с.) |


 15.3.1. МЕХАНОРЕЦЕПТОРЫ КОЖИ
15.3.1. МЕХАНОРЕЦЕПТОРЫ КОЖИ
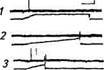


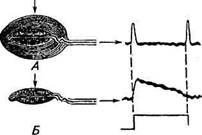 Рис. 15.8. Электрические ответы тактильного тельца Пачини на ступенчатообразное механическое раздражение:
Рис. 15.8. Электрические ответы тактильного тельца Пачини на ступенчатообразное механическое раздражение:
 до максимального значения, а затем в обоих случаях начинает уменьшаться до полного исчезновения. Таким образом, соотношение носит куполообразный характер (см. рис. 15.9, В).
до максимального значения, а затем в обоих случаях начинает уменьшаться до полного исчезновения. Таким образом, соотношение носит куполообразный характер (см. рис. 15.9, В).



