Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Частная физиология всасывания в кишечникеСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Всасывание воды и электролитов
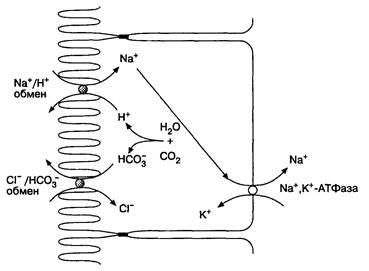
Содержимое тонкой кишки становится изоосмотичным прежде всего за счет двунаправленного перемещения как воды, так и электролитов. Выравнивание осмолярности обычно происходит в двенадцатиперстной кишке, поскольку объем химуса, поступающего туда одномоментно при нормальной работе желудка, небольшой. Если желудок опорожняется ненормально и в двенадцатиперстную кишку поступают большие объемы гиперосмотичного химуса, то в этом случае изоосмотичность в двенадцатиперстной кишке не может быть достигнута. Поскольку вода пассивно проходит в сторону гиперосмотичности, жидкость выходит из плазмы через ворсинки и поступает в просвет кишки. Значительный выход жидкости может привести к гиповолемии и вегетативным реакциям в виде тахикардии и потливости, что является признаками демпинг-синдрома. Главным механизмом абсорбции воды, электролитов и многих органических молекул является Na+,K+-ATФaзa, локализованная на базолатеральных участках мембран энтероцитов. Этот механизм транспорта требует затраты энергии АТФ и присутствия ионов магния для обмена трех ионов натрия, выходящих из клетки, на два иона калия, входящих в клетку. Поскольку суммарно клеткой теряются положительно заряженные ионы, электрический потенциал клетки относительно внеклеточной среды становится отрицательным. Na+,K+-ATФaзa создает градиент натрия, способствующий вхождению натрия обратно в клетку. Поэтому существует много веществ, транспортирующихся в клетку вместе с натрием: глюкоза, аминокислоты, ди- и трипептиды, соли желчных кислот. Для транспорта каждого из этих веществ необходим свой собственный белок-переносчик. Наличие ионов натрия значительно улучшает всасывание глюкозы; с другой стороны, глюкоза усиливает всасывание ионов натрия. На апикальном участке мембраны существуют также Nа+,Н+-антипорт и Сl~,НСО3–-антипорт (рис. 6-13). Благодаря этим белкам, Na+ и Сl– поступают в клетку, тогда как ионы водорода и бикарбонат выходят из клеток. Н+ и НСО3– образуются в клетке под действием карбоангидразы из СО2. Вода перемещается вслед за натрием пассивно: сначала в клетку, а затем из клетки в плазму крови.
Рис. 6-13. Всасывание воды и натрия в тонкой кишке. (По: Yamada Т., Alpers D. H., Owyang С., Powell D. W., Silver-stein F. E., eds. Textbook of Gastroentcrology, 2nd ed. Philadelphia: J.B.Lippincott, 1995; 1:333.)
Энтероциты также секретируют электролиты, в основном бикарбонат и ионы хлора. Секреция бикарбоната происходит преимущественно в проксимальном отделе тонкой кишки (но наблюдается и в других отделах) посредством Cl–, HСО3–-антипорта, но может осуществляться и с помощью иных механизмов. Секреция хлора связана с наличием на базолатеральном участке мембраны Na+,K+, Cl–-котранспортера (рис. 6-14), который переносит все три иона в клетку; это вторичный активный транспортный процесс. Как только хлор накапливается в клетке за счет действия Na,K,Cl и Сl, НСО3-транспортеров, происходит открытие хлорных каналов в апикальном участке мембраны, и хлор выходит из клетки в просвет кишки. Важность механизмов абсорбции и секреции хорошо выявляется при нарушении этих процессов. Токсин холерного вибриона стимулирует секрецию хлоридов и угнетает абсорбцию натрия и хлора в энтероцитах. При этом вода перемещается в просвет кишки вслед за натрием и вызывает сильную водную диарею. При пероральной регидратации необходимо введение растворов NaCl и глюкозы, поскольку Na+/глюкоза-котранспортер и Na+,K+-АТФаза не повреждаются холерным токсином и, следовательно, котранспорте? Na+/глюкoзa обеспечит сочетанное всасывание Na+ и глюкозы, а соответственно, и воды. Такое лечение значительно улучшает абсорбцию воды даже при холере.
Всасывание аминокислот
Переваривание белков в желудке происходит при превращении в кислой среде пепсиногена в пепсин (оптимальный рН 1—3). Пепсин расщепляет связи между ароматическими аминокислотами, соседствующими с карбоксильными аминокислотами. Пепсин инактивируется в щелочной среде. Этот этап переваривания белков отсутствует у больных после гастрэктомии, а также у тех, кто длительное время принимал ингибиторы Н+,К+-АТФазы, например омепразол. Расщепление пептидов пепсином прекращается после поступления химуса в тонкую кишку. В тонкой кишке полипептиды подвергаются дальнейшему расщеплению протеазами, имеющимися в соке поджелудочной железы и на поверхности микроворсинок энтероцитов. Основное расщепление пептидов происходит панкреатическими ферментами: трипсином, химотрипсином, эластазой и карбоксипептидазами А
Рис. 6-14. Секреция хлора в тонкой кишке. (По: Yamada Т., А1-pers D. H.,0wyang С., Powell D. W„ Silver-stein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995; 1:339.)
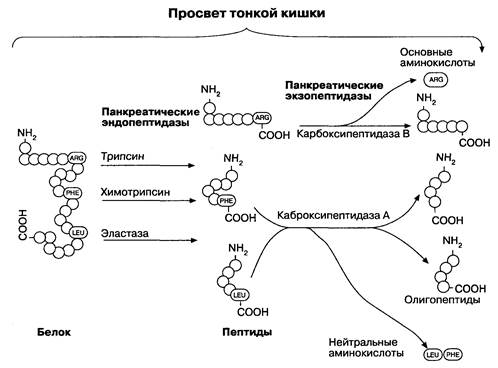
и В. Энтерокиназа переводит трипсиноген в трипсин, который затем активирует и другие протеазы. Трипсин расщепляет полипептидные цепочки в местах соединений основных аминокислот (лизина и аргинина), в то время как химотрипсин разрушает связи ароматических аминокислот (фенилаланина, тирозина, триптофана). Эластаза расщепляет связи алифатических пептидов (рис. 6-15). Эти три фермента являются эндопептидазами, т. к. они гидролизуют внутренние связи пептидов. Карбоксипептидазы А и В представляют собой экзопептидазы, поскольку отщепляют только концевые карбоксильные группы преимущественно нейтральных и основных аминокислот соответственно. При протеолизе, осуществляемом панкреатическими ферментами, происходит отщепление олигопептидов и некоторых свободных аминокислот. Микроворсинки энтероцитов имеют на своей поверхности эндопептидазы и экзопептидазы, которые расщепляют олигопептиды до аминокислот и ди-, трипептидов. Всасывание ди- и трипептидов осуществляется с помощью вторичного активного транспорта. Эти продукты затем расщепляются до аминокислот внутриклеточными пептидазами энтероцитов. Аминокислоты абсорбируются по принципу механизма котранспорта с натрием на апикальном участке мембраны. Последующая диффузия через базолатеральную мембрану происходит против градиента концентрации, и аминокислоты попадают в капиллярное сплетение ворсинок. Существует, как минимум, пять симпортов Na+/аминокислота, различающихся по типам переносимых аминокислот: нейтральный транспортер (переносящий нейтральные аминокислоты), основной (переносящий аргинин, лизин, гистидин), дикарбоксильный (транспортирующий глутамат, аспартат), гидрофоб-
Рис. 6-15. Переваривание белка в просвете кишки панкреатическими протеазами. (Но: Yamada Т.. Аlpers D. H., Owyang С., Powell D. W., Silverstein F. E., cds. Textbook of Gastroenterology, 2nd ed. Philadelphia:.;. B. Lippincott, 1995; 1: 457.)
ный (транспортирующий фенилаланин, метионин), и имино-транспортер (переносящий пролин, гидроксипролин). Описаны наследственные нарушения функций отдельных переносчиков аминокислот, приводящие к специфическим дефицитам аминокислот.
Всасывание углеводов
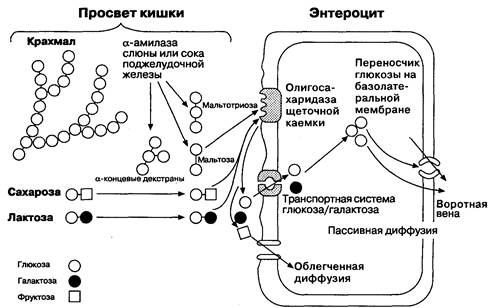
В кишечнике расщепляются и всасываются только те углеводы, на которые действуют специальные ферменты. Неперевариваемые углеводы, или пищевые волокна, не могут быть катаболизированы, поскольку для этого нет специальных ферментов. Однако возможен их катаболизм бактериями толстой кишки, что может вызывать образование газов. Углеводы пищи состоят из дисахаридов: сахарозы (обычный сахар) и лактозы (молочный сахар); моносахаридов: глюкозы и фруктозы; и растительных крахмалов: амилозы (длинных полимерных цепочек, состоящих из молекул глюкозы, соединенных al,4 связями) и амилопектина (другого полимера глюкозы, молекулы которой соединены a 1,4 и a 1,6 связями). Еще один углевод пищи — гликоген, является полимером глюкозы, молекулы которой соединены a 1,4 связями. Энтероцит не способен транспортировать углеводы размером больше, чем моносахарид. Поэтому большая часть углеводов должна расщепляться перед всасыванием. Амилазы слюны и поджелудочной железы гидролизуют преимущественно a1,4 связи глюкоза—глюкоза, но связи a1,6 и концевые связи a1,4 не расщепляются амилазой. Когда начинается переваривание пищи, амилаза слюны расщепляет a1,4-соединений амилозы и амилопектина, образуя a1,6-ветви a1,4-соединений полимеров глюкозы (так называемые концевые a-декстраны) (рис. 6-16). Кроме того, под действием амилазы слюны образуются ди- и триполимеры глюкозы, называемые соответственно мальтозой и мальтотриозой. Амилаза слюны инактивируется
Рис. 6-16. Переваривание и всасывание углеводов. (По: Kclley W. N., ed. Textbook of Internal Medicine, 2nd ed. Philadelphia:}. B. Lippincott, 1992:407.)
в желудке, т. к. оптимальный рН для ее активности составляет 6.7. Панкреатическая амилаза продолжает гидролиз углеводов до мальтозы, мальтотриозы и концевых a-декстранов в просвете тонкой кишки. Микроворсинки энтероцитов содержат ферменты, катаболизирующие олигосахариды и дисахариды до моносахаридов для их абсорбции. Глюкоамилаза или концевая a-декстраназа расщепляет а 1,4 связи на нерасщепленных концах олигосахаридов, которые образовались при расщеплении амилопектина амилазой. В результате этого образуются тетрасахариды с а1,6 связями, которые наиболее легко расщепляются. Сахаразно-изомальтазный комплекс имеет два каталитических участка: один с сахаразной активностью, а другой — с изомальтазной. Изомальтазный участок расщепляет а 1,4 связи и переводит тетрасахариды в мальтотриозу. Изомальтаза и сахараза отщепляют глюкозу от нередуцированных концов мальтозы, мальтотриозы и концевых а-декстранов; однако изомальтаза не может расщеплять сахарозу. Сахараза расщепляет дисахарид сахарозу до фруктозы и глюкозы. Кроме того, на микроворсинках энтероцитов также имеется лактаза, которая расщепляет лактозу до галактозы и глюкозы. После образования моносахаридов начинается их абсорбция. Глюкоза и галактоза транспортируются в энтероцит вместе с Na+ через Na+/глюкоза-транспортер; всасывание глюкозы значительно возрастает в присутствии натрия и нарушается в его отсутствие. Фруктоза, по-видимому, поступает в клетку через апикальный участок мембраны путем диффузии. Галактоза и глюкоза выходят через базолатеральный участок мембраны с помощью переносчиков; механизм выхода фруктозы из энтероцитов менее изучен. Моносахариды поступают через капиллярное сплетение ворсинок в воротную вену.
Всасывание жиров
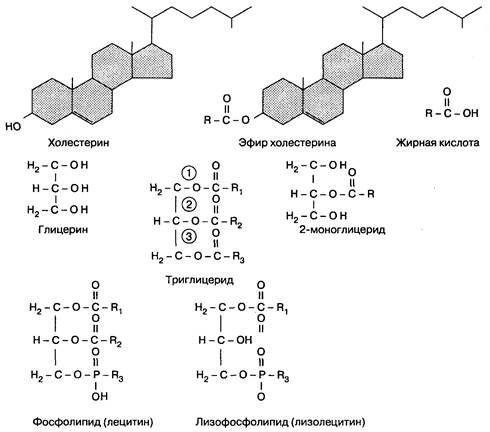
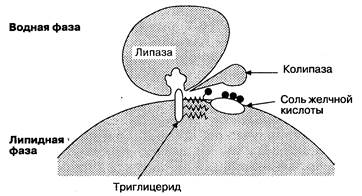
Жиры в пище состоят в основном из триглицеридов, фосфолипидов (лецитина) и холестерина (в виде эфиров) (рис. 6-17). Для полноценного переваривания и всасывания жиров необходимо сочетание нескольких факторов: нормальная работа печени и желчевыводящих путей, наличие панкреатических ферментов и щелочного рН, нормальное состояние энтероцитов, лимфатической системы кишечника и функциональной кишечно-печеночной циркуляции. Нарушение любого из этих компонентов приводит к нарушению всасывания жиров и стеаторее. В основном переваривание жиров происходит в тонкой кишке. Однако начальный процесс липолиза может проходить в желудке под действием желудочной липазы, вырабатываемой в дне желудка, при оптимальном значении рН 4—5. Липаза желудка расщепляет триглицериды до жирных кислот и диглицеридов. Она устойчива к воздействию пепсина, однако разрушается под действием протеаз поджелудочной железы в щелочной среде двенадцатиперстной кишки, ее активность снижается только под действием солей желчных кислот. Желудочная липаза имеет небольшое значение по сравнению с панкреатической липазой, хотя обладает некоторой активностью, особенно в антральном отделе, где при механическом перемешивании химуса образуются мельчайшие жировые капли, что повышает площадь поверхности для переваривания жиров. После попадания химуса в двенадцатиперстную кишку происходит дальнейший липолиз, включающий несколько последовательных стадий. Сначала триглицериды, холестерин, фосфолипиды и продукты расщепления липидов желудочной липазой сливаются в мицеллы под действием желчных кислот; мицеллы стабилизируются фосфолипидами и моноглицеридами в щелочной среде. Затем колипаза, секретируемая поджелудочной железой, воздействует на мицеллы и служит точкой приложения действия панкреатической липазы. В отсутствие колипазы панк
Рис. 6-17. Структуры липидов. (R = алкильпые цепи разной длины)
реатическая липаза обладает слабой липолитической активностью. Связывание колипазы с мицеллой улучшается в результате воздействия панкреатической фосфолипазы А2 (ФЛА2) на лецитин мицелл. В свою очередь, для активации ФЛА2 и образования лизолецитина и жирных кислот необходимо наличие солей желчных кислот и кальция. После гидролиза лецитина триглицериды мицелл становятся доступными для переваривания. Затем панкреатическая липаза прикрепляется к соединению колипаза—мицелла (рис. 6-18) и гидролизует 1 и 3-связи триглицеридов, образуя 2-моноглицерид и жирную кислоту. Оптимальный рН для панкреатической липазы составляет 6.0—6.5. Другой фермент — панкреатическая эстераза — гидролизует связи холестерина и жирорастворимых витаминов с эфирами жирной кислоты. Основными продуктами расщепления липидов под действием панкреатической липазы (ФЛА2) и эстеразы являются жирные кислоты, 2-моноглицериды, лизолецитин и холестерин (неэтерифицированный). Скорость поступления гидрофобных веществ в микроворсинки зависит от их солюбилизации в мицеллах в просвете кишки. Жирные кислоты, холестерин и моноглицериды поступают в энтероциты из мицелл путем пассивной диффузии (рис. 6-19); хотя жирные кислоты с длинной цепью могут переноситься и с помощью поверхностного связывающего протеина. Поскольку эти компоненты жирорастворимы и гораздо мельче, чем непереварен
Рис. 6-18. Взаимодействие панкреатической липазы и комплекса колнпаза-мицелла для гидролиза триглицеридов. (По: Sleisenger M. II., FordtranJ. S.,eds. Gastrointestinal Disease, 5th ed. Philadelphia: W. B. Saunders, 1993; 1:984.)
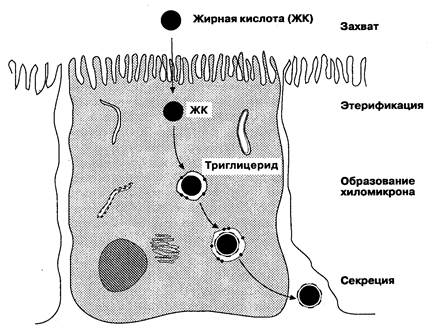
ные триглицериды и эфиры холестерина, они легко проходят через мембрану энтероцита. В клетке жирные кислоты с длинной цепью и холестерин переносятся связывающими протеинами в гидрофильной цитоплазме к эндоплазматическому ретикулуму (ЭР). Холестерин и жирорастворимые витамины переносятся стерольным белком-переносчиком к гладкому ЭР, где холестерин реэтерифицируется ацил-КоА-холестеринацилтрансферазой. Жирные кислоты с длинной цепью транспортируются через цитоплазму специальным белком, степень их поступления в шероховатый эндоплазматический ретикулум зависит от количества жиров в пище. При голодании (мало жиров) жирные кислоты соединяются с глицерин-3-фосфатом, образовавшимся при метаболизме глюкозы, для ресинтеза триглицеридов в шероховатом цитоплазматическом ретикулуме. При добавлении жиров в диету синте-
Рис. 6-19. Транспорт липидов в энтероците. (По: Isselbacher К. J. Biochemical reports of lipid malabsorption. Fed. Proc. 26: 1420, 1967;Johnson L. R., ed. Physiology of Gastrointestinal Tract, 2nd cd. New York: Raven Press, 1987: 1530.)
тический процесс переключается в гладкий ЭР, где образуются триглицериды из 2-моноглицеридов и жирных кислот. Лизолецитин, являющийся водорастворимым продуктом действия ФЛАз, может при наличии большого количества жиров преобразовываться в лецитин, взаимодействуя с жирными кислотами. При голодании лизоцетин, как и жирные кислоты, соединяется с глицерин-3-фосфатом с образованием конечной формы лецитина. После ресинтеза эфиров холестерина, триглицеридов и лецитина в ЭР они образуют липопротеины, соединяясь с аполипопротеинами, синтезируемыми в шероховатом эндоплазматическом ретикулуме энтероцитов. Липопротеины делят по размеру, по содержанию в них липидов и по типу апопротеинов, входящих в их состав. Хиломикроны и липопротеины очень низкой плотности имеют больший размер и состоят в основном из триглицеридов и жирорастворимых витаминов, тогда как липопротеины низкой плотности имеют меньший размер и содержат преимущественно этерифицированный холестерин. Липопротеины высокой плотности — самые маленькие по размеру и содержат главным образом фосфолипиды (лецитин). Дефицит аполипопротеинов нарушает транспорт липидов. При абеталипопротеинемии (состояние, при котором отсутствует необходимый для образования хиломикронов и липопротеинов очень низкой плотности апо-В-протеин) наблюдается переполнение энтероцитов жирами, нарушение всасывания жиров и неврологические расстройства, связанные с дефицитом витамина Е. Сформированные липопротеины выходят через базолатеральную мембрану в везикулах, далее они поступают в лимфатический сосуд. Жирные кислоты с короткой цепью, содержащей менее 12 атомов углерода, могут прямо поступать в систему воротной вены из энтероцитов без образования триглицеридов. Жирные кислоты с короткой цепью образуются под действием микроорганизмов из непереваренных в толстой кишке углеводов и являются важным источником энергии для колоноцитов.
|
||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 568; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.01 с.) |