Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Кислота и пепсин желудка. Измельчение и перемешивание пищиСодержание книги
Поиск на нашем сайте
В желудок пережеванная пища поступает через пищевод. Пищевые частицы подвергаются механической и химической обработке, превращаясь в гомогенную жидкую массу (химус), что улучшает процессы всасывания в тонкой кишке. Два главных процесса координируют пищеварение в желудке: секреция соляной кислоты и пепсиногена для химической обработки пищи, а также измельчение и перемешивание пищи, выполняемые специальными мышечными слоями. Кроме того, желудок вырабатывает внутренний фактор, являющийся обязательным кофактором для всасывания витамина b12. В G-клетках антрального отдела желудка образуется гастрин (пептид, стимулирующий секрецию кислоты).
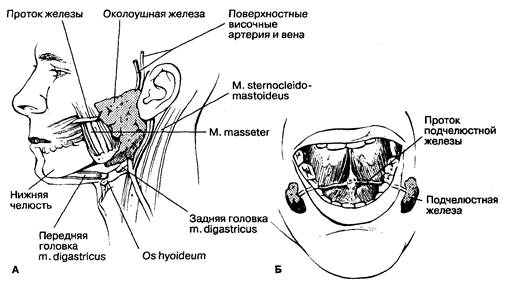
Рис. 6-1. Локализация околоушных (А) и подчелюстных (Б) слюнных желез. (По: Akesson E. J„ LoebJ.A,, Wilson-Pauwels L., eds. Thompson's Core Textbook of Anatomy, 2nd ed. Philadelphia: J. B. Lippincott, 1990: 335, 370.)
Рис. 6-2. Схема секреторных отделов подчелюстной железы. Секреторные элементы этой железы представляют собой либо серозно-мукоидные ацинусы (Ас), либо мукоидные протоки (МТ) с серозно-мукоидными полулуниями (SDL). Оба типа секреторных структур соединяются относительно большими промежуточными протоками (ID), полостными протоками (SD) и выводными протоками (ED). (По: John-son L. R., ed. Physiology of the Gastrointestinal Tract, 2nd cd. New York: Raven Press, 1987:774.)
Соляная кислота вырабатывается париетальными клетками, которые располагаются в перешейке трубчатых желез (рис. 6-3) вместе с добавочными клетками желудка (преимущественно в дне и теле желудка). Трубчатые железы желудка погружены в микроскопические ямки слизистой оболочки желудка и секретируют компоненты желудочного сока в просвет желудка. Обкладочные клетки образуют кислоту с помощью механизмов активного транспорта: Н+,К+-АТФаза на апикальной мембране клетки (рис. 6-4) выводит водородный ион из клетки, а ион калия поступает внутрь клетки. Париетальные клетки секретируют соляную кислоту с рН около 0.8, тогда как рН самой клетки составляет 7.2. На каждый секретируемый водородный ион образуется гидроксильный ион (ОН–), который сразу же реагирует с СО2 (катализируется карбоангидразой) с выделением бикарбоната и Н2О. Внутриклеточный бикарбонат на базолатеральном участке мембраны обменивается клеткой на внеклеточный Сl–, что увеличивает содержание внутриклеточного хлора, который транспортируется к апикальному участку мембраны и секретируется вместе с водородом, образуя НС1. Желудок выделяет около 2 л жидкости в сутки. Хотя кислота не является абсолютно необходимой для переваривания пищи, она улучшает переваривание белков и всасывание железа, а также защищает от патогенных микроорганизмов, попадающих в организм с пищей.
Рис. 6-3. Трубчатая железа желудка. (По: ItoS., Winchester R.J. The final structure of the gastric mucosa in the bat. J. Cell. Biol. 16: 541,1963; Yamada Т., Alpers D. H., Owyang C„ Powell D. W., Silvcrstein F. E„ eds. Textbook of Castroenterology, 2nd ed. Philadelphia:.!. B. Lippincott, 1995; 1: 297.)
Стимуляция секреции кислоты осуществляется посредством как нервных, так и гуморальных механизмов (рис. 6-5) и традиционно делится на три фазы. Данное деление на фазы связано не с самими механизмами стимуляции, а с источниками этой стимуляции или ингибирования секреции. Сложнорефлекторная фаза секреции желудочного сока вызывается видом, запахом и вкусом пищи и происходит через влияние блуждающего нерва на париетальные клетки. Желудочная фаза секреции осуществляется механическим растяжением желудка пищей, которое воспринимается специальными рецепторами растяжения в стенке желудка и реализуется через рефлекторную дугу, включающую блуждающий нерв. На секрецию в данной фазе влияют такие специфические компоненты пищи, как пептиды, аминокислоты, кофеин, этанол, кальций, которые стимулируют выработку гастрина — сильнейшего гуморального стимулятора секреции соляной кислоты. Кишечная фаза секреции желудочного сока связана с растяжением тонкой кишки химусом, а также с цирку-' пирующими в крови аминокислотами.
Рис. 6-4. Транспорт ионов в париетальной клетке желудка. Апикальный участок мембраны содержит Н+,К+-АТФазпый насос и каналы для транспорта К+ и Cl–. Базолатеральный отдел мембраны имеет насосы для К+, белки, обменивающие Сl–/НСО3– и Na+/H+, а также Nа+,K+-АТФазный механизм, которые поддерживают клеточный гомеостаз в состоянии покоя и при стимуляции секреции. (По: Yamada Т., Alpers D. В., Owyang С, Роwell D. W., Silverstein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995; 1:312.)
В основании трубчатых желез в теле и дне желудка располагаются главные клетки, секретирующие пепсиноген. Пепсиноген накапливается в зимогенных гранулах и высвобождается в просвет желудка под действием стимуляции блуждающим нервом (ацетилхолин) и, возможно, под действием пептидных гормонов, таких как гастрин и холецистокинин. В кислой среде пепсиноген аутокаталитически превращается в пепсин, обладающий в данной среде протеолитической активностью. Под влиянием пепсина происходит начальный этап переваривания белков в желудке, особенно разрушение коллагена. Пептиды, образующиеся при расщеплении белков пепсином, стимулируют выработку гастрина и холецистокинина, что является важным звеном в координации регуляции переваривания, необходимого для последующей абсорбции. Кроме того, кислый химус, поступая в тонкую кишку, стимулирует выработку не только холецистокинина, но также и секретина — гормона, способствующего образованию желчи и поджелудочного сока, богатых бикарбонатами. Липаза желудка, вырабатываемая клетками дна желудка, не играет существенной роли в катаболизме пищевых жиров. Желудок также выполняет функции депонирования и перемешивания пищи. Стенка желудка имеет три слоя мышц: наружный — продольный, средний — циркулярный и внутренний — косой. Циркулярный слой неравномерен: в дистальном отделе тела желудка и в антральной его части имеет большую толщину по сравнению с проксимальным отделом желудка. С помощью этих мышц желудок удерживает пищу. В момент, когда происходит механическая стимуляция глотки или пищевой комок растягивает пищевод, проксимальные отделы желудка (тело и дно) расслабляются для приема пищи (объемная релаксация). Этот процесс также регулируется блуждающим нервом. Растяжение желудка большими объемами пищи стимулирует перистальтические сокращения антрального отдела и проталкивание
Рис. 6-5. Регуляция секреции соляной кислоты в желудке. Показана основная (лиганд-рецепторная) регуляция выработки НС1 обкладочными клетками. D-клетки, соматостатиновые клетки; G-клетки, гастриновые клетки. (По: Feldman M. Acid and gastrin secretion in duodenal ulcer disease. Regul. Pcpt. Lett. 1: 1, 1989. Yamada Т., Alpers D. H., Owyang C., Powell D. W„ Silverstcin F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995; 1: 308.)
пищи к привратнику и двенадцатиперстной кишке. Проглоченные твердые частицы измельчаются до размеров меньше 1 мм, что увеличивает соотношение площадь—масса пищевых частиц и облегчает действие ферментов в тонкой кишке, приводя к более эффективному перевариванию. В желудке происходит избирательная задержка крупных частиц (желудочное "просеивание") и их измельчение, что предотвращает нарушение абсорбции питательных веществ. Измельчение и просеивание в желудке осуществляется за счет сокращения антрального отдела, с силой проталкивающего химус в направлении привратника, и столь же сильного обратного движения химуса при закрытии привратника. Эти движения разбивают крупные частицы химуса, если они не проходят в привратник, и позволяют мелким частицам (< 1 мм) пройти в двенадцатиперстную кишку. Пища, богатая питательными веществами, задерживается в желудке для более тщательной обработки и лучшего всасывания в тонкой кишке, что регулируется кишечно-желудочным рефлекторным механизмом. Углеводные компоненты пищи поступают в двенадцатиперстную кишку первыми, за ними следуют белки и затем жиры. Гиперосмолярная пища дольше задерживается в желудке благодаря механизму ингибирующей обратной связи после начала поступления ее в кишечник. Жидкая пища выходит из желудка пропорционально ее объему, этот механизм называется кинетикой "первого порядка". В целом жидкость выходит из желудка раньше твердой пищи, эвакуация которой происходит неравномерно: имеется начальный период задержки, затем — продолжительная линейная фаза, после которой наступает стадия очень медленного выхода. Линейная фаза выхода твердой пищи не зависит от ее объема и называется кинетикой "нулевого порядка". Привратник, антральный отдел и двенадцатиперстная кишка функционируют при опорожнении желудка как единый комплекс. Сокращения происходят последовательно от антрального отдела к привратнику и двенадцатиперстной кишке. Даже в случаях резекции привратника или пилоропластики процесс опустошения желудка сохраняется нормальным. Денервация проксимального отдела желудка ускоряет выход жидкости и не влияет на продвижение твердой пищи; хотя денервация антрального отдела с денервацией проксимального отдела ускоряет выход жидкости из желудка и замедляет выход твердой пищи.
Секреция желчи
Желчь продуцируется гепатоцитами и секретируется в кишечник в объеме около 500 мл в сутки. Желчь содержит соли желчных кислот, желчные пигменты, холестерин и другие липиды, а также щелочную фосфатазу. Желчные кислоты и их натриевые и калиевые соли необходимы для всасывания жиров. Эти соли конъюгируют с таурином или глицином, что повышает их гидрофильность и стабильность в тонкой кишке; по структуре они близки к холестерину. Гепатоциты вырабатывают холевую и хенодезоксихолевую кислоты, являющиеся первичными желчными кислотами. Под влиянием бактерий в тонкой кишке они модифицируются во вторичные желчные кислоты: дезоксихолевую, литохолевую и урсодезоксихолевую. Когда желчные кислоты через желчный проток и сфинктер Одди попадают в двенадцатиперстную кишку, они смешиваются с перевариваемыми липидами и жирорастворимыми витаминами, образуя при этом мицеллы, т. е. водорастворимые комплексы, из которых липиды легко абсорбируются. Образование мицелл стабилизируется в просвете кишечника в присутствии фосфолипидов и моноглицеридов, которые снижают их поверхностное натяжение. Мицеллы участвуют в эмульгировании жиров и увеличивают площадь поверхности для гидролиза, а также подготавливают жиры для всасывания в кишечнике. Желчные кислоты сами являются регуляторами выработки желчи в печени. Они реабсорбируются в тонкой кишке 4—15 раз в сутки вторичным активным транспортом (с электролитами) и попадают в систему воротной вены для рециркуляции. Как только желчные кислоты поступают в печень, по механизму отрицательной обратной связи происходит угнетение синтеза новых желчных кислот. Процесс кишечно-печеночной циркуляции желчных кислот представлен на Рис. 6-6. Без такой циркуляции наблюдается нарушение всасывания жиров, поскольку печень не в состоянии обеспечить достаточный синтез новых желчных кислот в количестве, соответствующем поступающим в кишечник липидам. В норме ежесуточная потеря желчных кислот с калом незначительна, и, следовательно, относительно невелик ежесуточный синтез желчных кислот в печени. Клетки желчного протока, как и клетки слюнных протоков, изменяют состав секрета за счет добавления в него бикарбоната и воды, поэтому окончательная желчь имеет щелочную реакцию и изоосмолярна плазме крови. Это обеспечивает нейтрализацию кислого химуса желудка. Желчный пузырь концентрирует желчь, вырабатываемую печенью, и выделяет ее в желчный проток и двенадцатиперстную кишку. Этот процесс регулируется холецистокинином. Клетки желчного протока увеличивают секрецию воды и бикарбоната под действием секретина. И секретин, и холецистокинин вырабатываются клетками глубоких отделов слизистой оболочки проксимального отдела тонкой кишки. Эти гормоны обладают синергическим действием на секрецию желчи и сока поджелудочной железы.
Рис. 6-6. Кишечно-печеночная циркуляция желчных кислот (ЖК) у здорового человека. (По: Carey M. С., Cahalane M. J. Enterohepatic circulation. In: Arias l.M.,JakobyW. В..Popper H., Schachter D., Shafritz D. A., eds. The Liver: Biology and Pathology, 2nd cd. New York: Raven Press, 1988:576; Yamada Т., Alpers D. H., Owyang C., Powell D. W., Silverstein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995; 1: 395.)
Панкреатическая секреция
Объем секрета поджелудочной железы составляет 1500 мл в сутки. Он выделяется в тонкую кишку и содержит ферменты, гидролизующие белки, жиры и крахмал. Основная регуляция секреции осуществляется гормонами. Холецистокинин стимулирует секрецию ферментов, а секретин, прежде всего, секрецию бикарбонатов. Существует также регуляция секреции через блуждающий нерв. Общая секреция поджелудочной железы зависит от стимулирующих и ингибирующих факторов. Пептидные гормоны — панкреатический полипептид, глюкагон и соматостатин — преимущественно угнетают секрецию поджелудочной железы. Панкреатический полипептид выделяется островками поджелудочной железы после стимуляции блуждающим нервом, ингибируя, таким образом, панкреатическую секрецию и секрецию желчи. Глюкагон угнетает панкреатическую секрецию в условиях гипергликемии. Панкреатическая секреция аналогично желудочной может быть разделена на три фазы. Сложнорефлекторная фаза секреции происходит в ответ на вкус и запах пищи и осуществляется через блуждающий нерв. Желудочная фаза — это стимуляция панкреатической секреции, осуществляемая при растяжении желудка также посредством блуждающего нерва. Наиболее сильная — кишечная фаза, в которую стимуляция секреции происходит, в основном, при растяжении кишки. Клетки протоков поджелудочной железы изменяют состав поджелудочного сока за счет выделения в него бикарбоната и воды. Для всасывания питательных веществ очень важны ферменты поджелудочного сока, выделяющиеся через панкреатический проток и сфинктер Одди. Такие протеазы, как трипсиноген, химотрипсиноген, прокарбоксипептидазы А и В и проэластаза выделяются из поджелудочной железы в неактивном виде. Трипсиноген превращается в двенадцатиперстной кишке в трипсин под действием энтерокиназы — фермента, вырабатываемого слизистой оболочкой двенадцатиперстной кишки (рис. 6-7). Трипсин, в свою очередь, переводит другие протеазы в активные формы химотрипсина, карбоксипептидаз А и В и эластазы. Он может также образовываться в результате самоактивации, т. е. перехода трипсиногена в трипсин. Секреция ферментов в неактивном виде предохраняет поджелудочную железу от самопереваривания, и если трипсиноген превращается в трипсин в поджелудочной железе, то возникает выраженное воспаление. Поджелудочная железа вырабатывает и ингибитор трипсина: его максимальная активность проявляется при рН, равном 3—7, но и она очень мала по сравнению с общей протеолитической активностью. Панкреатическая амилаза расщепляет полисахариды до олигосахаридов, липаза
Рис. 6-7. Активация панкреатических протеолитических ферментов. Эптерокиназа (энтеропептидаза) играет основную роль в переводе трипсиногена в трипсин. Трипсин, в свою очередь, активирует не только трипсиноген, но и другие предшественники' протеаз. (По: Sleisenger M. H., FordtranJ. S., eds. Gastrointestinal Disease, 5th cd. Philadelphia: W. B. Saunders, 1993; 1: 994.)
расщепляет эмульгированные триглицериды и эстераза — эфиры холестерина. Эти три последних фермента секретируются уже в активной форме, в отличие от протеаз, которые выделяются в виде неактивных предшественников. Колипаза, также секретирующаяся поджелудочной железой, является необходимым кофактором для действия липазы на триглицериды, так как нарушает взаимодействие между триглицеридами и желчными солями в мицеллах, и тем самым облегчает действие липазы на триглицериды.
Всасывание в кишечнике
|
||||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 287; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.128.168.219 (0.01 с.) |