Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Статева або репродуктивна системаСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Основна функція репродуктивної системи - розмноження, тобто притаманна всім організмам властивість відтворення собі подібних, що забезпечує безперервність життя. В основі всіх форм розмноження організмів, що мають клітинну будову, лежить поділ клітини. Основні способи розмноження - нестатевий, вегетативний і статевий. Виникнення статевого розмноження на основі більш примітивного нестатевого було прогресивним фактором еволюції, так як привело до підвищення спадкової мінливості й, відповідно, до прискорення темпів еволюції. Хордові майже усі розмножуються статевим шляхом. В абсолютній більшості вони роздільностатеві тварини.
Гермафродитами є лише личинкохордові. У зв'язку з цим у фенотипі більшості хордових розрізняють внутрішні та зовнішні статеві ознаки, які звичайно визначає генотип, але пізніше на їх прояв помітний вплив справляє гормональна система. До статевих органів належать статеві залози - гонади (сім'яники та яєчники), статеві протоки (сім'япроводи та яйцепроводи), копулятивні органи й так звані додаткові утворення. У гонадах формуються статеві клітини - гамети (яйцеклітини й сперматозоони), які виводяться з організму через яйцепроводи та сім'япроводи. Лише в деяких тварин гамети виводяться через протоки органів виділення. До додаткових утворень належать різні залози: у самців вони виділяють секрет, що служить фізіологічним середовищем для сперматозоонів і підсилює їх активність; у самок -речовини, з яких утворюються яйцеві оболонки. До додаткових органів належать також сім'яні сумки й сім'яприймачі - резервуари, де певний час зберігається сперма. У самок живородних тварин до статевих органів відносять і всі пристосування для виношування зародків. У губок і деяких війчастих червів гонад немає, їх статеві клітини розсіяні в паренхімі й виводяться через розриви стінки тіла або через ротовий отвір. У кишковопорожнинних статеві органи представлені невеликими гонадами в певних частинах тіла. Більшість плоских червів мають гермафродитну статеву систему з різними додатковими органами. Первиннопорожнинні черви роздільностатеві з порівняно простою статевою системою. У кільчастих червів, молюсків, ехіурид, моховаток та плечоногих статеві залози виникають у стінках целома; статевими протоками служать канали - целомодукти, не зв'язані з гонадами. Статеві продукти спочатку потрапляють у целомічну порожнину, потім у лійку целомодукта, який відкривається назовні. У членистоногих гонади являють собою похідні стінок вторинної порожнини тіла, а їх протоки - похідні целомодуктів. Серед хордових найбільш примітивними є статеві залози безчерепних (ланцетників). Вони представлені невеликими парними утворами, які тісно прилягають до стінки атріальної порожнини й не мають протоків. Статеві залози хребетних розвиваються у вигляді парного набрякання стінки порожнини тіла на боках спинної брижі між нею і нефростомами первинної нирки (мал. 1).
Мал. 1. Розвиток гонади та вивідних проток у самця стерляді. Чорним позначено первинні, крапками вторинні канальці мезонефроса (за Машковцевим): 1 - гонада; 2,3 - первинні й вторинні канальці; 4 - первиппонир-кова протока; 5 - первинна нирка; 6 — мезонефричний яйцепровід; 7 - переднирка.
Цей парний зачаток складається з поздовжньої складки потовщеного епітелію з внутрішньою масою сполучної тканини і має назву статевих або генітальних складок. Потім статеві складки вдаються глибше в порожнину тіла, відокремлюються від стінки тіла й стають, таким чином, підвішеними на брижі всередині порожнини тіла. У генітальні складки мігрують первинні статеві клітини, які виникають, як правило, за межами гонади на більш ранніх стадіях розвитку організму, і вростають шнури мезенхімних клітин з мезонефроса, які беруть участь в утворенні строми гонади. Епітелій гонади стає циліндричним, клітини його можуть перетворюватись у статеві, внаслідок чого він дістає назву зародкового епітелію. У риб статеві складки виникають по всій довжині порожнини тіла й цілком перетворюються в гонади. У земноводних вони закладаються так же, але перетворюється в гонаду лише середня частина складки. В амніот статеві складки з'являються тільки в ділянці бокової поверхні передньої половини мезонефроса, зверненої до мезентерію. У процесі розвитку гонади в обох статей проходять індиферентну (біпотенціальну) стадію розвитку, після якої починається їх статеве диференціювання. Яєчники (ovarium) формуються в основному з коркового шару гонади, сім'яники (testiculus) - з мозкового. Сім'яники в більшості тварин парні. Серед безхребетних вони найбільш просто побудовані в кишкопорожнинних і являють собою скупчення статевих клітин. Сперматозоони виводяться назовні шляхом розриву стінки тіла (гідроїдні поліпи) або через кишковосудинну систему й далі через ротовий отвір (сцифоїдні, коралові поліпи). Уперше від сім'яників відходять сім'япроводи в круглих червів. Серед нижчих хордових в апендикулярій сім'яник один і позбавлений протоків; у сальп - 2 з протоками; у асцидій від 1 до 150 пар сім'яників, від яких відходять протоки. У безчерепних сім'яники численні, позбавлені протоків. У більшості хребетних сім'яники - парні органи, симетрично розміщені в черевній порожнині, мають добре розвинені протоки. У самців нижчих хребетних зберігається ще до певної міри фолікулярна будова і в сім'яниках, проте окремі фолікули набирають вигляду справжніх міхурців і вступають у зв'язок з канальцями, що сполучають сім'яник з первинною ниркою. У вищих хребетних міхурці видовжуються в сім'яні трубочки, які вже сполучаються з сім'явиносними канальцями. Стінки сім'яних міхурців та сім'яних трубочок мають, крім первинних статевих клітин, також індиферентні, що служать для живлення статевих елементів під час їх розвитку.
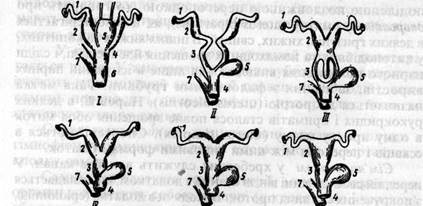
Статеві протоки. П оходження Під час ембріонального розвитку в деяких хребетних первиннониркова протока розщеплюється вздовж на два канали, з яких один зберігає зв'язок з первинною ниркою й називається вольфовим, а другий відокремлюється від неї і називається мюлеровим. У вищих хребетних мюлерів канал виникає як новоутворення в поздовжньому потовщенні стінки первиннониркової протоки. Передній кінець мюлерового каналу зростається з лійкою пронефроса й утворює яйцепровід (oviducti), який відкривається в порожнину тіла широкою миготливою лійкою, а другим кінцем впадає в клоаку. Можна вважати, що передні канальці переднирки служили спочатку не тільки для виділення сечі, але й для виведення жіночих статевих продуктів, а потім спеціалізувалися на цій функції, і разом з тим відокремилась також відповідна частина вивідної протоки (мал.93). У хрящових і кісткових ганоїдів, а також у костистих риб яйцепровід має інше походження: дві-три лійки пронефроса зростаються між собою і сполучаються з поздовжнім каналом нирки. Останній впадає в первиннониркову протоку. У вищих риб місце впадання відсувається назад, таким чином і цей яйцепровід відокремлюється від первиннониркової протоки. Яйцепроводи можуть потім диференціюватись на різні відділи. Так, у хрящових риб у ділянці передньої частини нирки кожен яйцепровід у статевозрілих особин утворює розширення - шкаралупову залозу, де яйце покривається спочатку драглистою білковою оболонкою, а поверх неї - майже в усіх яйцекладних видів - щільною рогоподібною оболонкою. Розширена задня частина яйцепровода називається маткою. Правий і лівий яйцепроводи відкриваються в клоаку самостійними отворами. У плазунів і птахів диференціація мюлерового каналу на відділи подібна, однак у птахів правий яйцепровід редукується. Запліднення яйця відбувається у верхній ділянці яйцепроводу. Виділення секреторних залоз середньої ділянки яйцепроводу утворюють навколо яйцеклітин (жовтка) білкову оболонку, а із секрету клітин стінок нижньої частини яйцепроводу (матки) формуються зовнішні яйцеві оболонки (підшкаралупова й шкаралупова). Для ссавців характерний процес поступового зростання яйцепроводів, який веде до утворення більш чи менш значних непарних відділів (мал. 2)
Мал. 2. Схема еволюції яйцепроводів у ссавців (за Холодковським): І-клоакові;II— нижчі сумчасті;III-вищі сумчасті (кенгуру); IV'— плацентарні з подвійною маткою; V — плацентарні з дворогою маткою; VI - плацентарні з простою маткою; 1 - фалопісва труба; 2 - матка; 3 - піхва; 4 — сечостатевий синус; 5 - сечовий міхур; 6 — клоака; 7 - пряма кишка.
У клоакових ссавців обидва яйцепроводи ще цілком розділені й самостійно впадають у сечостатевий синус. Задня частина кожного яйцепроводу розширена й утворює так звану матку, стінка якої виділяє рогоподібну шкаралупу яйця. Матки відкриваються в сечостатевий синус клоаки. У сумчастих нижні ділянки яйцепроводів перетворюються в довгі піхви, які нерідко зростаються своїми передніми кінцями. Від зрослого переднього кінця піхов назад напрямлений сліпий мішок (мал. 94), який іноді ззаду зростається з сечостатевим синусом і проривається в нього. Таким способом розвивається так звана третя піхва. Решта яйцепроводів диференціюється на власне яйцепроводи або фалопісві труби, що відкриваються в порожнину тіла широкою лійкою з бахромчастим краєм, і матки (uterus), в яких розвивається зародок. У плацентарних ссавців задні кінці мюлерових проток зливаються в непарну піхву (vagina). Якщо кожна матка відкривається в піхву самостійним отвором, її називають подвійною (uterus duplex) з парним отвором у піхву (більшість гризунів, слони та ін.) (мал. 94). Якщо матки зливаються тільки ззаду, тим часом як передня частина лишається поділеною поздовжньою перегородкою, то говорять про двороздільну матку (uterus bipartitus), як це спостерігається в деяких гризунів, хижих, свиней. В інших хижих та копитних, у китоподібних та комахоїдних зрощення йде ще далі, і сліди парного походження виявляються лише в існуванні парних виростів, зв'язаних з фалопієвими трубами. Така матка називається дворогою (uterus bicornis). Нарешті, в деяких рукокрилих і приматів сталося повне зрощення обох маток в одну просту матку (uterus simplex). Спостерігаються в ссавців і переходи між цими типовими формами маток. Сім'япроводом у хребетних служить вольфів канал. У передній своїй ділянці він зв'язаний з додатком, що складається з покручених сім'яних проток. Виходячи з додатка (epididimis), сім'япровід у вигляді рівного каналу іде до впадання в сечостатевий синус. Останній витягнутий у вигляді каналу, який називається сечостатевим (canalis urogenitalis) і має добре розвинену мускулатуру. Сечостатевий канал продовжується в канал копулятивного органа. У всіх хрящових риб та земноводних вольфів канал служить одночасно сім'япроводом і сечоводом. У костистих риб сім'япровід являє собою новоутворення, не зв'язане з мезонефросом. Парні сім'япроводи відкриваються назовні спільним сечостатевим або статевим отвором. У плазунів і птахів вольфові канали служать тільки сім'япроводами. У зв'язку зі статевими протоками в ссавців розвиваються різні додаткові залози, особливо характерні для самців. Біля основи статевого члена лежить передміхурова залоза або простата (prostata), секрет якої утворює рідку частину сперми. По боках сечостатевого каналу знаходяться парні куперові залози, що мають поперечносмугасту мускульну оболонку й беруть участь в утворенні ейякулята. Зі статевим членом зв'язані також препуціальні залози, які виділяють пахучий секрет, що сприяє зустрічі особин протилежної статі. Гомологи цих залоз є і в присінку в самки, де вони відомі під назвою бартолінісвих.
ЗОВНІШНІ СТАТЕВІ ОРГАНИ Копулятивні органи хребетних можуть мати різне походження. Так, у самців хрящових риб вони являють собою видозмінені базалії черевних плавців (птеригонодії), які мають на своїй внутрішній поверхні жолобок, по якому стікає сперма з клоаки самця. У костистих риб копулятивний апарат спостерігається рідко. У живородних коропозубих таким апаратом служить видозмінений анальний плавець (гоноподій), яким самець переносить сперматофор і прикріпляє його до статевого отвору самки. Серед земноводних копулятивні органи є тільки в гімнофіон, у самців яких довга клоака може вивертатися в клоаку самки. Самці всіх плазунів, крім гатерії, мають копулятивні органи, що являють собою непарні (у крокодилів і черепах), або парні вирости задньої стінки клоаки, які при заплідненні вивертаються назовні. Копулятивні органи у вигляді непарного виросту стінки клоаки є лише в деяких видів птахів - страусів, тінаму, гусеподібних. В інших запліднення відбувається при притисканні зовнішнього отвору клоаки самця до клоаки самки. Найскладніше розвинені копулятивні органи ссавців. Це непарний орган, що розвивається в зародка в передній стінці клоаки у вигляді невеликого спочатку виступу - клоакового горбка. У передній його ділянці розвивається статевий горбок, на нижній поверхні якого утворюється поздовжній статевий жолобок, обмежований по боках статевими складками. Навколо всіх цих частин розвивається кільцеподібна складка - статевий валик. Подальший розвиток цих індиферентних частин статевого апарату відбувається по-різному в особин різної статі - у самця статеві валики зростаються між собою по середній лінії й утворюють стінки мошонки, а в самки приматів - так звані великі губи. У самця статевий горбок росте і дає статевий член (penis); у самки він слабо розвивається й перетворюється в клітор. Статеві складки по краях статевої борозенки дають у самки багатьох ссавців малі губи, а сечостатевий синус - присінок піхви. У самця краї статевих складок зростаються над сечостатевим синусом та над статевою борозенкою і так розвивається сечостатевий канал. У клоакових ссавців копулятивний орган знаходиться всередині клоаки, оточений складкою слизової оболонки, і може висуватися назовні та втягатися назад у клоаку під час парування. У живородних ссавців клоака зникає, і між сечостатевим синусом та каналом копулятивного органа настає постійне й неперервне сполучення, так що він служить не тільки для виведення сперми, а й для сечовипускання. Статевий член розміщається тепер ззовні, але оточується складкою шкіри, що утворює його піхву.У багатьох ссавців в пенісі є спеціальна кісточка, або бакулюм (гризуни, рукокрилі, нижчі примати тощо). Отже, хребетні, як правило, роздільностатеві. Статеві залози частіше парні. Яєчники мають більш-менш помітну зернисту структуру. Сім'яники відрізняються гладкою поверхнею. Для анамній характерне зовнішнє запліднення, однак у хрящових і деяких кісткових риб, хвостатих земноводних запліднення внутрішнє. Яйця анамній здатні розвиватись тільки у водному (рідко - в дуже вологому) середовищі. Лише в деяких груп спостерігається яйцеживородіння або живородіння. З яйця вилупляється личинка, яка веде водний спосіб життя. Для амніот характерна ускладнена будова яйця (збільшення кількості жовтка й білка, утворення щільних захисних зовнішніх оболонок), внутрішнє запліднення, здатність яйця розвиватись тільки в повітряному середовищі. Це досягнуто зміною ходу ембріогенезу (формування зародкових оболонок - серозної, амніотичної, алантоїса). Розвиток - без метаморфозу. Усі ці особливості яйця, ембріонального та постембріонального розвитку були частиною тих важливих еволюційних набутків, які дозволили амніотам, на відміну від анамній, вийти з водного середовища й заселити всі ділянки суші. Отже, у становленні та вдосконаленні репродуктивної системи мали місце: диференціація клітин на статеві й соматичні; розвиток статевих залоз, статевих шляхів і копулятивних органів; спочатку гермофродитизм, а далі роздільностатевість з менш або більш вираженим статевим диморфізмом; спочатку зовнішнє, а далі внутрішнє запліднення; виникнення пристосувань для розвитку плода в матці та живлення його за рахунок організму матері через плаценту.
|
||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 556; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.41 (0.011 с.) |