Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Методи порівняльної анатоміїСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Дані порівняльної анатомії тварин разом з даними палеонтології та ембріології служать основним доказом еволюції, пояснюють виникнення рудиментів та атавізмів. Методи, якими користуються морфологи при вивченні і сучасних форм, і особин на ранніх етапах розвитку їх в онтогенезі (ембріологія) та при вивченні викопних решток цих тварин (палеозоологія), спрямовані завжди на пошуки гомологій і для встановлення на їх основі порівняльно- анатомічних рядів. Останні можуть більш-менш точно відображати справжні філогенетичні зв'язки і, отже, можуть бути використані для створення справжньої, не штучної, системи тваринного світу. При цьому зоологи користуються практично всіма методиками, які на різних рівнях живого можуть бути використані морфологами для встановлення гомологій. На рівні цілого організму використовують методи препарування на макро- та макро-мікроскопічному рівнях з використанням оптичних приладів малого збільшення (лупи, стереоскопічні лупи тощо). Для вивчення будови тканини різних органів використовують гістологічні методи (виготовлення тонких зрізів тканин на мікротомах та фарбування їх різними фарбами, гістохімічні методики та використання світлової мікроскопії). На клітинному рівні використовують електронну мікроскопію. При дослідженні кровопостачання проводять наливку судин різними контрастними масами з наступною ренгенографією. На трупах цей метод доповнюється вживанням корозійних засобів. Дослідження живих організмів проводять з використанням ренгенографії, томографії, ультразвукової діагностики, ендоскопії та інше. Після описування різних структур організму приступають до порівняння їх між собою (своєрідна філософія в біології). Кожен із цих методів сам по собі може багато дати для вивчення історії розвитку тваринного світу шляхом встановлення гомологій, а при вживанні їх у комплексі (так званий метод потрійного паралелізму) значення їх ще більше зростає. Проте усі вони мають свої недоліки та обмеження. Справа в тому, що перенести результати вивчення морфології сучасних форм на викопні досить проблематично, бо сучасні види тварин складають лише близько 2% від усіх видів, які будь-коли існували на Землі, вони багато в чому відрізняються у звуязку з появою адаптивних змін. Тоді, можливо, опорою в цій справі може бути палеонтологія, яка насправді зробила вагомий внесок в пізнання історичності зоології? На жаль, палеозоолог працює лише з рештками. Це рештки скелетів, до того ж далеко не усіх форм, які колись існували на Землі. Тобто багато перехідних форм нам недоступні, а можливо, й ніколи не будуть виявлені. Має свої недоліки й ембріологія, хоча у свій час Мюллер та Геккель покладали на неї великі надії, сформульовані коротко так: "Ембріогенез - це стисла, скорочена форма філогенезу". Справа в тому, що на ембріональному рівні в історичному вимірі відбуваються значні адаптивні зміни, які були названі архалаксісами, девіаціями та анаболіями в залежності від того, на якій стадії розвитку ембріона вони виникають. Враховуючи це Сєверцов і розробив цілу науку - філембріогенез. Тому О.М.Сєверцов (1939) вважав, що в майбутньому зоологи для встановлення філогенетичної системи тваринного світу будуть використовувати крім чисто морфологічних методів ще й знання з фізіології, етології, екології. Він дав назву такому підходу "еволюційна морфологія" і вважав, що за нею майбутнє. Лекція 2. Шкіра та її похідні у хордових тварин (2 год.) Логіка виходу 1. Будова шкіри у водних тварин. 2. Шкірні покриви наземних хребетних. 3. Шкірні залози хордових. 4. Рогові утвори шкіри хребетних. 5. Шкірні скостеніння. Рекомендована література: 1, 2, 3, 4, 5 ПОКРИВИ ТІЛА ТА ЇХ ПОХІДНІ Шкірні покриви тварин - це система зовнішніх органів, яка має безпосередній зв'язок з оточуючим середовищем, у першу чергу реагує на зміни в умовах довкілля. Покриви тіла виконують роль захисту від шкідливих впливів, функцію сприймання подразнень, беруть участь в обміні речовин, синтезі вітаміну Д, терморегуляції, є додатковим органом дихання і виділення. Зв'язок тварини з зовнішнім середовищем здійснюється за допомогою ектодерми. Тому сприймання подразнення, чутливість і захист тіла від шкідливих впливів - основні функції ектодерми, які в нижчих тварин і залишаються цілком за нею. Спеціалізація окремих клітин і цілих ділянок ектодермального покриву для сприймання тих чи інших подразнень приводить до високого диференціювання. Отже, у шкірі розрізняють поверхневий шар - епідерміс і підстилаючий його мезодермальний шар - сполучнотканинну частину шкіри, або коріум. У всіх хордових тварин завжди можна чітко розрізнити обидва шари шкіри: верхній, епідермальний - епідерміс і нижній, мезодермальний - коріум. В індивідуальному розвитку первинний шкірний покрив - шар ектодерми - дає зовнішній шар шкіри, тобто епідерміс з його похідними (залози, роговий шар, органи чуття, пігменти тощо). Власне шкіра, або коріум - сполучнотканинний шар шкірних покривів - розвивається за рахунок мезодерми від шкірного листка соміта. У всіх хордових тварин завжди можна чітко розрізнити обидва шари шкіри: верхній, епідермальний - епідерміс і нижній, мезодермальний - коріум. В індивідуальному розвитку первинний шкірний покрив - шар ектодерми - дає зовнішній шар шкіри, тобто епідерміс з його похідними (залози, роговий шар, органи чуття, пігменти тощо). Власне шкіра, або коріум - сполучнотканинний шар шкірних покривів - розвивається за рахунок мезодерми від шкірного листка соміта. Починаючи від круглоротих у всіх хребетних наявне ускладнення шкірних покривів. Порівняльна характеристика будови та функцій шкіри в амніот і анамній
Починаючи з круглоротих у всіх первинноводних хребетних, або анамній, будова шкіри характеризується загальними рисами: 1) епідерміс багатошаровий з великою кількістю різноманітних слизових та зернистих клітин, що функціонують як одноклітинні залози; 2) коріум з щільними рядами колагенових волокон, які формують поздовжні та вертикальні шари, що правильно чергуються.
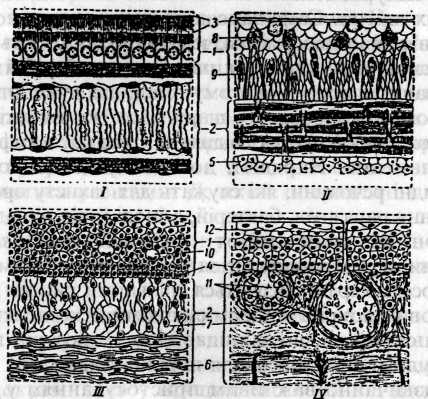
У шкірі круглоротих і риб є залозисті клітини різного типу: бокалоподібні, що відкриваються на поверхню епідермісу; кулясті з зернистим вмістом; колбоподібні, що сидять на ніжці, яка доходить до основи епідермісу. Останні виділяють рідину в міжклітинні простори, а не безпосередньо на поверхню тіла (мал.4).
Мал 4. Розріз через шкіру хордових (за Нілом і Рейду). І - ланцетник; II -мінога; III - акула; IV - ропуха. 1 - епідерміс; 2 - коріум; 3 - кутикула; 4 - мальпігіїв шар клітин; 5 - підшкірна сполучна тканина; 6 - щільна і 7 - пухкі сполучні тканини; 8 - зернисті клітини; 9 - колбоподібні клітини; 10- слизові клітини; Типових багатоклітинних шкірних залоз у круглоротих і риб немає. Залозисті заглибини епідермісу при основі голок та шипів служать у деяких риб як отрутовидільні залози. Складні утвори залозистого характеру являють собою й органи свічення деяких глибоководних риб. У личинок земноводних є багато шкірних залозистих клітин, однак у дорослих вони зникають і заміняються багатоклітинними залозами. Вони розвиваються за рахунок клітин основного шару епідермісу, які заглиблюються в сполучнотканинну частину шкіри і дають початок групі кулястих клітин, що лежать під епідермісом. Приблизно на початку метаморфозу всередині такого зачатка залози з'являється порожнина, яка продовжується у вигляді вузького вивідного каналу, що проходить через епідерміс на його зовнішню поверхню. Клітини розміщаються в один шар по стінці залози. Багатоклітинна залоза дорослої тварини має вигляд простого міхура, стінка якого складається з одного шару залозистих клітин. Сама залоза лежить у коріумі й вузькою шийкою зв'язана з епідермісом, через який проходить її вивідний канал. Основна функція шкірних залоз амфібій полягає в підтримуванні шкіри у вологому стані, що дуже істотно через слабкий розвиток її рогового шару. Зволоження шкіри є також умовою можливості шкірного дихання, яке має в амфібій досить велике значення. Нарешті, до складу цих залоз входять бактерицидні речовини, які служать для захисту організму від проникнення паразитів, бактерій, а іноді й від нападу ворогів. Шкірні покриви амніот - плазунів, птахів і ссавців, незважаючи на різкі відмінності за зовнішніми ознаками (лусочки, пір'я, волосся), характеризуються спільними рисами будови. Слизовий покрив епідермісу анамній заміщається сухим роговим покривом, що захищає тіло амніот від висихання. Ця перебудова епідермісу для первинноназемних хребетних стала надзвичайно важливим пристосуванням для життя на суші. Ороговіння шкірних покривів відбувається завдяки появі в цитоплазмі епідермальних клітин зернинок рогової речовини - кератогіаліну, які поступово заповнюють усю клітину, після чого вона відмирає. Епідерміс амніот поділяється на мальпігіїв, або ростковий шар живих клітин і роговий шар з відмираючими клітинами. Коріум амніот складається зі сплетіння сполучнотканинних волокон (колагенових та еластичних). В амніот (крім ссавців) різко скорочується кількість шкірних залоз. Так, у плазунів як виняток зустрічаються лише окремі великі залози, що виконують особливі функції. Їх виділення відіграють роль хімічних сигналів для привертання уваги особин протилежної статі, мічення територій. У крокодилів є пара мускусних залоз по боках нижньої щелепи; у черепах подібні залози лежать на місці сполучення карапакса і пластрона; у ящірок - стегнові пори. У птахів шкірних залоз також немає, за винятком однієї парної залози, що виконує спеціальні функції. Це - куприкова залоза, що знаходиться на хвостових хребцях. Вона утворена з численних трубок, що радіально сходяться до вивідного каналу, і виділяє маслянистий секрет, коли птах надавлює на залозу дзьобом. Цим секретом птахи змащують пір'я, що сприяє його еластичності, створює гідрофобний шар. Жирові виділення куприкової залози під дією сонячного світла перетворюються у вітамін Д, який потрапляє в організм птахів. Шкіра ссавців на відміну від інших амніот багата шкірними залозами: потовими, сальними, молочними, пахучими (мал.5). Усі вони є похідними епідермісу й лише вторинно глибоко занурені в коріум. Наявність залоз у шкірі ссавців при відсутності їх у плазунів і птахів свідчить про те, що предки ссавців відокремились у процесі еволюції від дуже давніх плазунів, які зберегли спільні риси з викопними земноводними. У шкірі ссавців є багатоклітинні залози обох основних типів - трубчасті й альвеолярні. До перших належать потові залози, що мають вигляд довгої трубки, кінець якої нерідко згорнутий клубочком. Стінки потової залози складаються з одношарового епітелію, вкритого зовні шаром гладеньких мускульних волокон. Потові залози відкриваються абобезпосередньо на поверхню епідермісу, або у волосяну сумку. Епітеліальні клітини цих залоз секретують піт, що на 97 - 99 % складається з води, у якій розчинені сечовина й креатин, жирні кислоти й солі. Отже, з потом виділяються продукти розпаду, однак основна функція потових залоз терморегуляційна. Потовиділення регулюється тепловими центрами головного й спинного мозку. Потові залози добре розвинені в приматів і копитних, дуже мало їх у зайцеподібних, гризунів, хижих, відсутні в китоподібних, ящерів, сиренових, ластоногих.
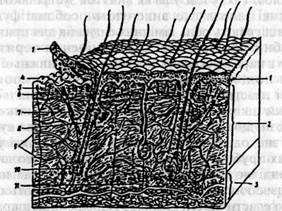
Мал.5. Розріз через шкіру людини: 1- епідерміс; 2 - коріум; 3 - підшкірна жирова клітковина; 4 - дермальні сосочки; 5 - нервові закінчення; 6 - капіляри; 7-м 'яз, що рухає волос; 8 - сальна залоза; 9 - кровоносна судина; 10 - корінь волоса; 11 - потова залоза.
До типу альвеолярних належать сальні залози. Це гронуваті залози, що складаються з кількох лопатей, зв'язаних з сумками волосся, у які вони виділяють свій жирний секрет. Розвиваються сальні залози як вирости від піхов волосся. Вони не мають мускульного покриву. Епітелій їх багатошаровий, і його клітини при утворенні секрету зазнають жирового переродження, руйнуються й виштовхуються. Секрет сальних залоз змазує поверхню шкіри й волосся, сприяючи збереженню еластичності, а також запобігає проникненню бактерій, є водовідштовхуючим покривом. Молочні залози типових ссавців за своєю будовою є також складними гронуватими утворами й тривалий час вважались продуктом перетворення сальних залоз, бо секрет їх багатий на жир. Проте тепер доведено, що молочні залози є наслідком видозміни трубчастих потових залоз. У яйцекладних ссавців молочні залози зберігають трубчасту будову, мають мускульний покрив із гладеньких м'язових волокон і відкриваються у волосяні сумки на особливому залозистому полі самиці: у качкодзьоба - на череві, у єхидни - у виводковій сумці. Малята злизують крапельки молока з волосся. У живородних ссавців залозисте поле обмежується невеликою ділянкою покривів, випнутою у вигляді соска. Самі молочні залози мають складнішу будову й належать до альвеолярного типу. Проте в сумчастих, якщо й не на самих залозах, то на їх протоках зберігається гладенький мускульний покрив, який доводить існування генетичного зв'язку цих залоз з молочними залозами клоакових ссавців, а також і з потовими залозами. Треба додати, що й секрети їх, як і секрет останніх, є результатом життєдіяльності клітин, а не наслідком їх руйнування, як у сальних залозах. Це дозволяє твердити, що молочні залози всіх ссавців являють собою перетворені потові залози. Пахучі залози ссавців являють собою перетворені потові, рідше сальні, інколи поєднання тих і других. Такими є параанальні залози багатьох хижаків, особливо куничих, мускусні залози кабарги, бобрів, ондатри, передочні залози багатьох парнокопитних, копитні залози кіз тощо. Пахучий секрет цих залоз служить перш за все для мічення територій і для видового розпізнавання. Рідше різкий (скунси) пахучий секрет використовується для самозахисту. Сукупність запахів секретів пахучих, сальних і потових залоз дозволяє тваринам відрізняти особин свого та інших видів, полегшує зустріч самців і самок тощо. У більшості видів риб у шкірі утворюються захисні утвори - лусочки. У шкірі хрящових риб угворюється плакоїдна луска. Вона складається з широкої основи, що лежить у коріумі, і шипа, вістря якого виступає через епідерміс назовні. Усередині луски є порожнина, наповнена багатим на кровоносні судини м'якушем - пульпою. Плакоідна луска складається з дентину, утвореного клітинами коріуму. Вістря шипа покрите емаллю, утвореною клітинами базального шару епідермісу. В акул така луска більш чи менш рівномірно покриває все тіло; у скатів у шкірі розкидані великі лусочки, між якими безладно розсіяні дрібні, а в деяких видів луска редукується. Плакоїдна луска більш складної форми й більших розмірів розміщена на щелепах і перетворена в зуби. Доведено, що зуби всіх хребетних побудовані за одним типом і являють собою видозмінені плакоїдні лусочки предків, тобто є їх гомологами. Це свідчить про спільне походження всіх хребетних від акулоподібних предків з плакоїдною лускою. Вивчення шліфів лусок викопних риб, а також ембріонального розвитку лусок сучасних кісткових риб показує, що зачатки плакоїдних лусок є складовою частиною складних ганоїдних і космоїдних лусок. У розвитку лусок кісткових риб можна визначити З етапи: 1) ускладнення кісткових лусок шляхом зростання окремих плакоїдних лусок і перетворення дентину в космін - космоїдні луски кистеперих риб, викопних дводишних, а також акантодій із панцирних риб; 2) подальше ускладнення шляхом розвитку нового поверхневого шару ганоїну та глибинного шару ізопедину при перетворенні космоїдної луски в ганоїдну в сучасних кісткових ганоїдів, багатоперих та у викопних палеонісок; 3) спрощення кісткової луски шляхом редукції всіх шарів, за винятком тонкого кісткового шару в циклоїдних та ктеноїдних лусках у всіх сучасних костистих риб, а також у мульної риби з кісткових ганоїдів. Отже, усі види луски риб генетично зв'язані один з одним. Крім лусок у більш глибоких шарах коріуму утворюються шкірні кістки. У найдавніших риб вони були дуже численні, порівняно невеликі, мали не цілком установлене розміщення. З'явившись у шкірі, ці кістки у вищестоячих форм занурюються глибше і вступають у тісний зв'язок з частинами внутрішнього скелета - черепом та плечовим поясом, утворюючи на ньому так звані покривні кістки. Вторинні (накладні) кістки не зв'язані з первинними (вихідними) функціями хрящового скелета. Такі кістки утворюють зяброву кришку, яка має велике значення для активізації дихання. Вторинний плечовий пояс здійснює зв'язок первинного пояса з черепом, а вторинні щелепи забезпечують можливість споживати різний корм. У наземних хребетних рідко зберігаються сліди покриву з шкірної луски. Панцир з луски циклоїдного типу був у багатьох викопних амфібій іноді на всьому тілі або лише на череві. Із сучасних земноводних залишки шкірної луски збереглися в гімнофіон у вигляді маленьких кісткових пластинок, які лежать глибоко в шкірі. Але якщо в сучасних наземних хребетних, як правило, загальний лускатий покрив зник, то в них усіх збереглись його похідні - рогові утвори. Основне значення рогового покриву наземних хребетних полягає в захисті шкіри від висихання та пошкодження. Значне потовщення та видозміна рогового шару приводить у вищих хребетних до утворення особливих органів, що служать для захисту тіла від механічних пошкоджень і виконують функцію зовнішнього скелета, або до розвитку особливого покриву, який зменшує тепловіддачу. Утвори першого роду особливо поширені в плазунів (панцир), а другого - у птахів і ссавців (пір'я та волосся). У рептилій роговий шар потовщується невеликими ділянками, між якими він залишається тоншим, завдяки чому зберігається рухливість тварини. Таким способом розвивається найпримітивніша форма луски - горбкувата (гекони, хамелеони, гатерія). Луска у вигляді пластинок, які черепицеподібно налягають одна на одну заднім краєм, характерна для ящірок і змій. Іноді луска перетворюється в шипи та різні складні вирости. На нерухомих частинах тіла, наприклад, на голові у всіх плазунів, а також на спинному й черевному щитах черепах, розвиваються плоскі рогові пластинки - щитки. В інших хребетних рогова луска звичайно не утворює загального покриву тіла: у птахів вона покриває тільки нижні частини ніг, а у ссавців зустрічається як виняток, особливо на хвості (бобри, щури та ін.). У птахів типові рогові луски є лише на ногах, а на тілі розвивається пір'я, яке має різну будову. Будова лускоподібних пір'їн на крилах пінгвінів, вторинно спрощених у зв'язку з пристосуванням до пірнання, і на ногах страусів дозволяє зробити висновок про походження пір'я з лускоподібних утворень. Це підтверджується ембріональним розвитком (мал.6). Ембріональний зачаток пера гомологічний зачатку рогової луски. Молодий зачаток пера з'являється як скупчення мезодермальних клітин, що трохи піднімають епідерміс у вигляді горба, який потім починає рости назад (мал.6). На цих стадіях розвитку зачаток пера цілком схожий на зачаток луски. Потім шкіра, що оточує основу зачатка, поглиблюється, утворюючи піхву пера, а сам зачаток складається з центральної маси мезодерми (м'якуш пера), оточеної товстим епідермісом. В епідермісі утворюються поздовжні потовщення, які заглиблюються в м'якуш пера і потім цілком відділяються від поверхневого шару, що утворює чохлик на молодому пері. Самі потовщення роговіють і дають борідки ембріонального пуху, які вільно розпадаються, коли чохлик скидається. У сосочку розвиваються кровоносні судини, які живлять ростуче перо. Ембріональний пух заміняється дефінітивним пером, яке виростає під ним на тому ж сосочку й поступово його виштовхує. Своєрідним утвором шкірних покривів ссавців є волосся. На відміну від рогових лусок і пір'я, волосся має чисто епідермальне походження і тільки в цибулині шерстин є сполучнотканинний сосочок. Лише в деяких ссавців (китоподібні, слони, носороги, бегемоти) волосяного покриву на тілі практично немає. Дослідження показали, що під час ембріогенезу зародки цих тварин і людини на деякий час покриваються волоссям, що свідчить про вторинний характер його втрати в дорослому стані. Наявність рогових лусок у деяких ссавців (бобри, нутрія, хохуля, неповнозубі) одночасно з волоссям показує, що волосся розвивалось самостійно між роговими лусками, лише пізніше замінило їх. Розвиток волосся, на відміну від розвитку луски та пір'я, починається в епідермісі. Деяка група клітин набирає вищої циліндричної форми й починає швидко розмножуватись. Епідермальний зачаток заходить глибоко в мезодермальну частину шкіри, а потім у масі його клітин починається диференціювання: зовнішні шари дають початок зовнішній і внутрішній піхвам і сальним залозам, а глибока маса клітин перетворюється в зачаток власне волосини. Знизу зачаток заходить у мезодермальний сосочок, що живить його. Клітини зачатка волоса починають роговіти, і в міру того, як волосина наростає знизу за рахунок діяльності цибулини, кінець її пробивається через піхву назовні. Волосяний покрив виконує термоізолюючу функцію, служить рецептором дотику, захищає шкіру від пошкоджень і паразитів, поліпшує аеро- й гідродинамічні властивості тіла, забезпечує видоспецифічність забарвлення. Крім такого загального покриву, як луска плазунів, пір'я птахів, волосся ссавців, у наземних хребетних спостерігаються ще місцеві потовщення рогового шару різного значення. Так, на щелепах черепах, птахів та клоакових ссавців є товстий роговий шар, що утворює дзьоб і заміняє відсутні в цих тварин зуби. На кінцях пальців в амфібій є деяке потовщення рогового шару, яке іноді має вигляд справжніх кігтів. У рептилій і птахів усі пальці озброєні кігтями, що охоплюють їх кінці у вигляді чохлика. Первинна функція кігтів - захист кінців пальців від пошкоджень; у подальшому кігті стають допоміжними органами при пересуванні тварин і виконують різні пов'язані з цим функції; нарешті, у ссавців вони іноді перетворені в органи активного захисту тварини й навіть у знаряддя нападу. Так у деяких ссавців є копита - товсті рогові утворення, що майже цілком охоплюють кінцеву фалангу й надійно захищають її. У багатьох приматів кігті перетворились у нігті, що прикривають кінці пальців тільки зверху, а знизу розвинена м'яка подушка, що підвищує роль пальців як органів дотику. За рахунок розростання рогового епітелію в порожнисторогих утворюються роги - товсті рогові чохли, що мають постійний характер. Кісткові вирости, на яких сидять роги, утворюються з участю особливого шкірного окостеніння, яке приростає до черепа. Роги оленів - кісткові утворення, похідні коріуму. Це провізорні органи. Отже, шкірні покриви хребетних - це система зовнішніх органів, яка має прямий зв'язок з оточуючим середовищем і в першу чергу реагує на зміни в умовах зовнішнього середовища. Основними функціями шкіри та її похідних є: захист організму від несприятливих зовнішніх впливів; участь в обміні речовин (водному, сольовому, газообміні); функції органа дотику; інколи участь у терморегуляції (птахи, ссавці); захист організму від проникнення хвороботворних агентів за допомогою механізмів фагоцитарної та біохімічної природи; сприймання механічних, температурних, больових впливів. Шкіра та її похідні визначають забарвлення тварин, виділяють пахучі, отруйні або поживні речовини, що служать сигналами, захистом або засобом вигодовування потомства. У літаючих тварин або плаваючих їх високі аеро- чи гідродинамічні властивості значною мірою обумовлені властивостями та активним функціонуванням шкіри та її похідних. Шкіра нерозривно зв'язана з життєдіяльністю всього організму. Різні порушення функцій органів і систем нерідко відображаються на стані шкірних покривів.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 592; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.135.209.231 (0.017 с.) |