Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Сейсмосенсорні органи (бічна лінія)Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Сейсмосенсорні органи або бічна лінія являють собою спеціалізовані шкірні органи чуття, характерні для круглоротих, риб та личинок земноводних. Вони формуються на ектодермальних потовщеннях шляхом клітинного диференціювання невеликих ділянок, що дають початок окремим відчуваючим горбкам. У круглоротих та амфібій ці органи являють собою потовщення ектодерми, які лежать на поверхні або в неглибоких ямках і представлені грушоподібними вторинночутливими клітинами та циліндричними опірними клітинами, що йдуть через увесь орган від його поверхні до основи. Чутливі клітини мають на своєму вільному кінці щетинку або джгутик, а в основі обплітаються кінцевими закінченнями бічної гілки блукаючого нерва. Ці органи розміщені в один -три повздовжніх ряди на боковій поверхні тіла і в декілька рядів, інколи у вигляді складної мережі на голові. У риб у товщі шкіри утворюються канали, що відкриваються назовні спеціальними отворами. У костистих риб ці отвори пронизують лусочки й покривні кістки. На стінках каналів розміщені окремі сенсори бічної лінії (мал. 3). Крім системи каналів у риб спостерігаються й окремі органи бічної лінії, розміщені на поверхні шкіри або заглиблені в ямки -невромасти. Більш складними спеціалізованими є мішечки ганоїдів та слизові канали селахій (ампули Лоренціні).
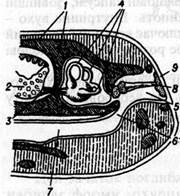
Мал. 3. Схема органа бічної лінії кісткової риби (за Гундричем): А - поздовжній розріз; Б - вигляд збоку. 1 - канал; 2 - зовнішні отвори каналу; 3 -луска; 4 -рецептори бічної лінії; 5 — бокова гілка блукаючого нерва; б — відгалуження нерва до органа бічної лінії. Функціональне значення органів бічної лінії полягає в сприйманні рухів та коливань води, завдяки чому тварини можуть орієнтуватись в потоках води, а сприймаючи хвилі, що йдуть від інших пливучих тварин, або відбиті хвилі власного пересування, можуть виявляти наближення хижака чи здобичі, пливти не натикаючись на підводні предмети і орієнтуватись у просторі. У наземних хребетних органи бічної лінії зникають. Органи слуху та рівноваги Органи слуху - це спеціалізовані органи чуття, що служать для сприймання та аналізу звукових коливань. У багатьох безхребетних, примітивних хордових вони сприймають не тільки звукові, а й механічні (вібраційні) коливання низькочастотного діапазону. Серед безхребетних справжні органи слуху розвинені тільки в комах. У більшості випадків вони представлені тимпанальними органами, а інколи й іншими утворами, чутливими до звукових коливань (хордотональні, джон-стонові органи). Тимпанальні органи зустрічаються в прямокрилих, метеликів, напівтвердокрилих. Розміщені вони,на ногах, черевці або грудях. Складаються з тонкої кутикулярної перетинки, зв'язаної з трахеями або повітряними порожнинами, інколи захищеними складкою скелета, і групи хордотональних сенсил, прикріплених або до перетинки, або до трахей. Усі типи таких органів слуху сприймають низькочастотні коливання; у деяких метеликів і прямокрилих тимпанальні органи сприймають ультразвукові коливання певних тварин, наприклад ехолокаційні сигнали рукокрилих. Органи слуху хордових як структурно, так і за походженням зв'язані з органами рівноваги. У водних хребетних такий комплексний орган утворений капсулами внутрішнього вуха. Як показує вивчення ембріонального розвитку, зачаток органа слуху гомологічний за походженням органам бічної лінії. Капсули внутрішнього вуха закладаються по боках голови зародка у вигляді парних потовщень ектодерми (мал. 4).
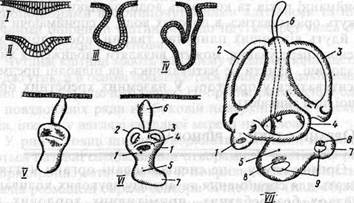
Мал. 4. Схема розвитку внутрішнього вуха хребетних: І- VII послідовні стадії (за Шмальгаузеном): 1 - овальний мішечок; 2-4 півколові канали; 5 - круглий мішечок; б - ендолімфатична протока; 7-лагена (початок завитки): 8 - слухові чутливі поля; 9 — отоліти.
Цей зачаток поглиблюється у вигляді ямки, що потім набирає форми міхурця, який спочатку сполучається з поверхнею каналом, а пізніше відшнуровується від ектодерми. Слуховий міхурець диференціюється на два відділи, відокремлені один від одного перехватом. З верхнього відділу формується овальний мішечок (utriculus) і з'єднані з ним три півколові канали, розміщені в трьох взаємно перпендикулярних площинах. Це - орган рівноваги (вестибулярний апарат). Нижній відділ перетворюється в круглий мішечок (sacculus), від якого відходить порожнистий виступ - лагена, яка у вищих хребетних перетворюється в довгий більш чи менш зігнутий канал завитка, в середині якого знаходиться власне орган слуху (кортіїв орган). З круглим мішечком зв'язаний довгий, зазвичай замкнутий ендолімфатичний канал - залишок сполучення порожнини первинного слухового міхурця з зовнішнім середовищем. Отже ускладнення слухового лабіринту в ряду хребетних відбувається так: 1) поступово відокремлюються півколові канали; 2) відокремлюється круглий мішечок (sacculus), який відділяється від овального мішечка (utriculus), у ньому розвиваються отоліти; 3) у наземних хребетних утворюється завитка (равлик), де формується складно збудований кортіїв орган. У птахів це лише зігнутий канал, у людини - 2,5 завитка, у гризунів їх біля 5. Орган слуху й рівноваги риб представлений тільки внутрішнім вухом, розміщеним в хрящовій капсулі, зовнішні стінки якої у кісткових риб костеніють. Внутрішнє вухо -власне перетинчастий лабіринт - включає круглий і овальний мішечки і з'єднані з останнім добре розвинені три півколові канали (мал.6, VII). Звуки на внутрішнє вухо поступають від веберового апарату (мал. 6).
Мал. 6. Схема органа слуху костистої риби (за Ромером, Парсонсом): 1 - внутрішнє вухо; 2 - ендолімфатичний мішок; 3 - перилімфа; 4 - кісточки веберового апарату; 5 - плавальний міхур; 6 - хребці. Цей апарат історично виник інакше. Для передачі коливань служить плавний міхур. У оселедцеподібних костистих риб від міхура відходить трубчастий відросток, який з'єднується з перетинчастим лабіринтом і передає коливання міхура ендолімфі. В іншої групи риб, до якої входять сомові, коропові, існує інший механізм звуко-передачі, а саме: парні відростки передніх хребців перетворились в окремі елементи- веберові кісточки. Вони з'єднуються в ланцюжки з двох боків і зв'язують плавальний міхур зі слуховим апаратом. Вони функціонують подібно слуховим кісточкам середнього вуха ссавців. Перехід з водного середовища, що добре передає всі механічні подразнення, в повітряне, де хвилі механічного подразнення швидко затухають, обумовив розвиток у всіх наземних хребетних нового відділу органа слуху - середнього вуха (мал.7), в якому розміщений апарат, що підсилює сприймання звукових коливань.
Мал.7. Схема поперечного розрізу голови жаби в ділянці вуха (за Григо-рі). 1— черепна коробка; 2 — довгастий мозок; 3 - слуховий нерв; 4 — пів-колові канали; 5 - порожнина середнього вуха; 6 - свстахісва труба; 7 — глотка; 8 — стремінце; 9 - барабанна перетинка. Порожнина середнього вуха утворилась з рудимента зябрової щілини, розміщеної між щелепною та під'язиковою дугами - бризкальця. Зовнішній отвір порожнини середнього вуха затягнутий тонкою та пружною барабанною перетинкою (membrana tympani). У порожнині знаходиться палочкоподібна кісточка - стремінце (stapes), яка одним кінцем впирається в барабанну перетинку, а другим - в овальне вікно, яке закрите меншою порівняно з барабанною перетинчастою перетинкою. Це й дозволяє підсилити слабі звукові коливання. Порівняльно-анатомічні та ембріологічні дослідження показали, що стремінце утворилось з підвіска - верхнього елемента під'язикової дуги, що стало можливим у зв'язку з розвитком аутостилії. Порожнина середнього вуха сполучається з задньою частиною ротової порожнини звуженим проходом -євстахієвою трубою (tuba Eustachii), яка утворилась з рудимента зябрової щілини, розміщеної між першою та другою зябровими дугами. Завдяки євстахієвій трубі (гомологу бризкальця) вирівнюється тиск, що запобігає розривам барабанної пертинки при сильних звуках. Серед наземних хребетних найпримітивніший орган слуху в амфібій. Однак поряд с системою (середнє і внутрішнє вухо), яка забезпечує слух у повітряному середовищі, в земноводних зберігаються механізми слуху у воді. Звукопровідність їх тіла, подібно до риб, близька до водного середовища, що дозволяє приймати звуки безпосередньо на мембрану овального вікна. Цьому сприяють поверхневі вени й ендолімфатична протока. Звуки у воді сприймаються так званим macula sacculi - гомологом звукорецептора риб, а звуки в повітрі, що передаються стремінцем з барабанної перетинки, сприймаються papilla neglesta amphibiorum - звуковим рецептором наземних хребетних. Існування двох звукових систем у земноводних дозволяє їм пристосуватись до життя у двох середовищах -водному й повітряному. У деяких безхвостих (кумки), у всіх хвостатих і безногих амфібій порожнина середнього вуха та барабанна перетинка вторинно редукуються. У воді ці тварини використовують описаний вище механізм звукосприймання, а можливості їх слуху в повітряному середовищі, мабуть, невеликі. Орган слуху плазунів схематично близький до органів слуху безхвостих земноводних. Равлик (лагена), що служить апаратом аналізу та кодування звукових сигналів, ще відсутній в більшості видів, являє собою мішкоподібний виріст. |Це відповідає відносно невеликій ролі слуху в житті плазунів. Вони сприймають звуки в діапазоні 20 - 6000 Гц, але найкраще чують тільки в діапазоні 60 - 200 Гц. У змій слух особливо слабо розвинений; вони не мають барабанної перетинки й сприймають переважно звуки, що розповсюджуються по землі або у воді (так званий сейсмічний слух). Передача звуків з субстрату на овальне вікно середнього вуха забезпечується квадратною та квадратно-виличною кістками. Порівняно обмежений слух відповідає їх слабим голосовим можливостям. Невисокі слухові здібності черепах, барабанна перетинка в і' яких товста, а слуховий прохід у деяких видів закритий потовщеною шкірою. Орган слуху птахів анатомічно подібний до органа слуху найбільш високоорганізованих плазунів (крокодилів), однак завдяки незначним перебудовам функціонально він не відрізняється від більш складного та диференційованого органа слуху ссавців. Внутрішнє вухо птахів відрізняється від внутрішнього вуха крокодилів лише кращим розвитком лагени та збільшенням кількості чутливих клітин. Збільшуються розміри середнього вуха; єдина слухова кісточка або стовп (columella). Стремінце має ускладнену форму, що збільшує її рухливість при коливанні барабанної перетинки, яка в птахів має круглу форму й великі розміри. Барабанна перетинка заглиблена нижче від рівня шкіри, до неї веде канал - зовнішній слуховий прохід, по краю якого в деяких видів (совиних) утворюється складка шкіри - зачаток зовнішнього вуха. Птахів з погано розвиненим слухом, мабуть, немає. Більшість видів сприймають звуки в діапазоні від ЗО до 20 тис. гц. Найбільш складний і досконалий орган слуху ссавців. Він представлений трьома відділами: зовнішнім, середнім і внутрішнім вухом (мал. 8).
Мал. 8. Схема будови органа слуху ссавця. 1 - зовнішній слуховий прохід; 2 - ендолімфатич-ний канал; 3 - кругле вікно; 4 - ковадло; 5 - молоточок; 6 - барабанна перетинка; 7 — слуховий нерв; 8 - є встахієва труба; 9 - нерв завитки; 10 - стремінце. Зовнішнє вухо (вушна раковина) та зовнішній слуховий прохід являють собою звукову антену - фільтр, здатну відбирати, визначати напрям надходження звуку й підсилювати важливі для виду звуки, послаблюючи зайві шуми. Таку ж роль виконує середнє вухо, в якому три слухові кісточки: молоточок (malleus), ковадло (incus), стремінце (stapes) утворюють важільну систему, що передає коливання з більшої за розмірами барабанної перетинки на меншу за площею мембрану овального вікна внутрішнього вуха, підсилюючи ці коливання. Кісточковий барабан, у якому розміщена порожнина середнього вуха, утворює ряд камер - резонаторів, що підсилюють біологічно важливі звуки. У деяких видів, особливо норників, вони інколи заповнені губчастою кістковою масою, що гасить паразитичні шуми. Внутрішнє вухо розміщене в кам'янистій частині вискової кістки й складається, як і в інших хребетних, з вестибулярного та слухового відділів. Максимального розвитку досягає завиток (равлик) - спірально зігнута перетинчаста трубка, яка лежить у кістковому футлярі й заповнена ендолімфою. У її центрі розміщена по всій довжині базальна мембрана, на якій впоперек натягнені фібрили (слухові струни). До них прилягають чутливі клітини кортієвого органа, що сприймають коливання слухових струн. Імпульси, сприйняті чутливими клітинами, передаються нейронам, аксони яких утворюють слуховий нерв. Такий механізм забезпечує тонкий аналіз частотного спектра й часової організації слухового сигналу, прийнятого зовнішнім вухом і переданого посиленим через середнє внутрішньому вуху. За широтою звукового діапазону ссавці переважають птахів, широко використовуючи як надзвукові (більше 20 кГц), так і низькі частоти. Отже, поступове вдосконалення вловлюючого та передаючого апарату (зовнішнє й середнє вухо) так само характеризує наземних хребетних, як і вдосконалювання власне сприймаючого звук апарату внутрішнього вуха.
Органи зору Органи сприймання світлових подразнень характерні для більшості безхребетних (за винятком губок) і всіх хребетних. У багатоклітинних тварин основний елемент органа зору - первинночутлива зорова клітина - фоторецептор. Сприймання світла здійснюється її периферичним кінцем (зовнішнім сегментом), який у хребетних має форму палички або колбочки. Найпростіші органи зору складаються з окремих світлочутливих клітин, розміщених серед епітеліальних клітин на поверхні тіла. Вони характерні для дощових червів, здатних розрізняти тільки темряву й світло. Ці клітини містять у собі особливе внутрішнє тіло, але не зв'язані з будь-якими скупченнями пігменту. Ускладнення органів сприймання світла в процесі еволюції відбувалося шляхом концентрації окремих світлочутливих клітин у скупчення й заглиблення їх під покриви тіла, створення пігментних екранів, а також світлозаломлюючих, акомодаційних, окорухових та захисних пристосувань. У деяких кишковопорожнинних та нижчих червів органи зору являють собою очні плями, розміщені в ектодермі, й складаються зі світлочутливих та пігментних клітин. Ускладнюючись, органи зору набувають частіше келихоподібної форми; інколи порожнина заповнена прозорою драглистою масою, що заломлює світло, - склистим тілом. У багатьох членистоногих (ракоподібних, комах) органи зору представлені фасетковими очима, що складаються з численних окремих очок - оматидіїв. Такі очі дозволяють сприймати форму предметів, пристосовані до бачення на близькій відстані. Парний орган зору хребетних до певної міри схожий на найдосконаліші органи зору безхребетних - очі головоногих молюсків. Однак при ближчому розгляді виявляється, що розміщення елементів ретини в них є зворотним, кришталик у хребетних розвивається в покривах, а не в передній стінці очного міхура, і тому схожість між цими органами основана лише на аналогії. На питання про походження органів зору хребетних проливає деяке світло факт світлочутливості самої центральної нервової системи в ланцетника. В останнього в передній частині нервової трубки є окремі світлочутливі клітини (очка Гессе), дуже схожі на очка війчастих червів. Світлочутлива клітина має на своєму внутрішньому кінці крайовий шар, який складається з великої кількості паличок і яким клітина заглиблюється в пігментну клітину, що має форму чаші. Зовнішній, обернений до світла, кінець клітини продовжується в нервовий відросток. Ці очка Гессе за своїм типом є оберненими органами зору, як і парні очі хребетних. У личинок асцидій є невелике, але трохи досконаліше очко, що розвивається також у стінках мозкового міхурця. Непарні очі (не більше двох) хребетних розміщаються по середній лінії на спинній поверхні голови. Ці органи розвиваються як порожнисті вирости покриття проміжного мозку, які поступово відшнуровуються й перетворюються в окремі міхурці, сполучені з мозком за допомогою ніжки. У більшості хребетних ці органи дають потім утвори залозистого характеру, але в круглоротих та в багатьох плазунів тут розвиваються справжні додаткові органи зору. Передня стінка очного міхурця стає прозорою, а задня перетворюється в сітківку, від клітин якої у напрямі до мозку розвиваються нервові відростки, що утворюють більш чи менш довгий нерв. У круглоротих розвиваються два таких міхурці: передній - це тім'яний орган або парієтальний, а задній лежить над першим безпосередньо під прозорими покривами і являє собою пінеальний орган. В інших хребетних пінеальний орган втрачає характер органа зору й перетворюється в залозу (glandula epiphisis cerebri). Тім'яний орган набуває іноді окоподібної будови або, принаймні, вигляду міхурця у вищих риб (дводишних, мульних), але звичайно він редукується. Тільки в деяких плазунів парієтальний орган має характер міхурчастого ока. Спинні очні міхурці хребетних належать до типу міхурчастих очей. Однак від очей безхребетних вони відрізняються своїм розвитком від центральної нервової системи. Можливо, що це - органи, парні за походженням, які вторинно змістились до середньої лінії. Судячи з існування тім'яного отвору в черепі багатьох викопних риб (кистепері) та стегоцефалів, можна припустити, що раніше ці органи мали більше поширення. Парні очі хребетних тварин закладаються на ранніх стадіях зародкового розвитку (мал.6). Одночасно з обособлениям проміжного мозку на його стінках утворюються бічні випини - очні міхурці. З ростом зародка очні міхурці відтісняються дедалі більше від мозку, їх стеблинки (ніжки) видовжуються й звужуються, а зовнішня частина стінки кожного міхура впинається всередину останнього й утворюються два очних келихи з подвійною стінкою.
Мал.9. Схема розвитку ока хребетних. I-IV- послідовні стадії (за Шмальгаузепом): 1 - проміжний мозок; 2 - очний міхур; З - ніжка очного міхура; 4 - кришталик; 5 - сітківка; 6 - пігментний шар; 7 — судинна оболонка; 8 - склера; 9 — рогівка.
Ектодерма над очними келихами утворює потовщення, яке заглиблюється під покрив назустріч очному келиху спочатку у вигляді ямки, а потім замкнутого міхурця, що являє собою зачаток кришталика. Він цілком відокремлюється від ектодерми й розміщається в отворі очного келиха; внутрішня, більш товста стінка очного келиха перетворюється в сітківку, а більш тонка зовнішня - в пігментний шар. Із мезодерми, що оточує очний келих, формується судинна оболонка, яка служить для живлення сітківки, і склера, що являє собою скелет ока. Ця скелетна оболонка продовжується й попереду кришталика, де вона зростається з покривами, утворює прозору рогівку. Так розвиваються найістотніші частини ока. Схема будови ока єдина у всіх класах хребетних, відмінності полягають лише в деталях будови, різних способах акомодації. Очі риб мають плоску рогівку й кулястий кришталик. Немає ні м'язів зіниці, ні війкового тіла, ні рухливих повік. Тонкий виріст судинної оболонки - серпоподібний відросток - пронизує сітківку й склисте тіло, прикріпляється своїм кінцевим здуттям до сумки кришталика. Цей відросток має гладенькі м'язові волокна, при скороченні яких кришталик переміщається всередину. З протилежного боку зв'язка служить антагоністом, який відтягує кришталик до попереднього місця. Тобто акомодація досягається переміщенням самого кришталика. У багатьох риб на внутрішній поверхні судинної оболонки утворюється дзеркальце (тапетум) - шар плоских клітин, заповнених кришталиками гуанину. Він відбиває світлові промені, які пройшли через сітківку, що посилює можливості бачення при слабій освітленості середовища. Порівняно з рибами в амфібій рогівка більш опукла, кришталик має форму двоопуклої лінзи з більш плоскою передньою поверхнею. У радужній оболонці та у війковому тілі є гладенька мускулатура. Акомодація досягається переміщенням кришталика за допомогою особливого м'яза війкового тіла. Під час метаморфозу утворюються рухливі повіки, які мають захисне значення. Органи зору наземних хребетних (амніот) краще пристосовані до роботи в повітряному середовищі, де кут заломлення світлових променів значно більший, ніж у воді. Вони захищені повіками; добре розвинені слізні залози. У птахів і плазунів від задньої стінки очного яблука в склисте тіло вдається своєрідний виріст, або гребінець (pecten), багатий кровоносними судинами. Він покращує живлення сітківки. Акомодація досягається завдяки кривизні рогівки і зміні кривизни кришталика за допомогою поперечносмугастого війкового тіла. На відміну від земноводних, аналіз і синтез зорових відчуттів відбувається не в сітківці, а переважно в зоровій корі проміжного мозку. У ссавців склера ока волокниста. Нерідко між склерою та судинною оболонкою є шар клітин з кришталиками, що відбивають світлові промені (tapetum). Це зумовлює "свічення" очей відбитим світлом (хижаки, копитні). Акомодація досягається шляхом зміни форми кришталика завдяки скороченню війкового м'яза. Останній не діє безпосередньо на кришталик. У спокійному стані кришталик: розтягнутий сумкою, в якій знаходиться, і має більш плоску форму (встановлення зору вдалечінь). При скороченні війкового м'яза послаблюється натяг сумки і кришталик стає опуклішим, як і в птахів. Райдужка відіграє роль діафрагми, регулюючи освітленість сітківки зміною величини зіниці. Отже, парні органи зору хребетних розвиваються як вирости з центральної нервової системи, тобто виникли не безпосередньо в покривах, як очі безхребетних, а шляхом диференціювання нервової системи. Що таке диференціювання дійсно можливе, доводить факт знаходження світлочутливих клітин у нервовій трубці ланцетника та личинки асцидій. Коли та як саме виникли очі хребетних, точно не відомо, але загальна картина більш-менш ясна. Можна припустити, що в межах такої пластинки настала потім диференціація, яка привела до утворення очних плям, а згодом і очних ямок. Коли нервова пластинка опустилась глибше й замкнулась у трубку, очні ямки набрали вигляду виступів на цій нервовій трубці. На користь такого погляду говорять факти з історії ембріонального розвитку. Трохи інакше виглядає ця картина на основі порявняльно-анатомічних даних. Бовері та Сєвєрцов вважали, що органи зору хребетних утворились філогенетично трохи пізніше, на стадії, коли нервова пластинка вже замкнулась у трубку. У ланцетника центральна нервова система ще має вигляд борозенки, на дні якої розміщені світлочутливі вічка. У предків хребетних зі збільшенням розмірів тіла, зменшенням прозорості тканин і утворенням трубки мозок потрапляє в менш вигідні умови для сприймання світлових подразнень. В еволюції хребетних ці несприятливі моменти були усунені, бо деякі ділянки мозку з великою кількістю світлочутливих елементів почали випинатися у вигляді міхурів, наближаючись, таким чином, до покривів, які в цих місцях зберегли чи навіть посилили свою прозорість. Потовщення ектодерми утворило світлозаломлююче тіло - кришталик, набуття якого було значним кроком уперед, бо зробило можливим образний зір. З наближенням очних міхурів до покривів кривизна кришталика повинна була збільшуватись. Через те, що очні міхури розвивались від мозку, найбільше значення для сприймання світлових подразнень мала близька до покривів зовнішня стінка очних міхурів. Опукла форма цієї стінки була несприятлива для образного зору, бо кришталик відкидав усередину сферично вгнуте зображення, вгнутість якого збільшувалась разом із ростом кривизни кришталика, тобто з наближенням очних міхурів до покривів. Тому передня (зовнішня) стінка очних міхурів вдавлювалась всередину, і так розвинулись очні келихи, внутрішня (передня) стінка яких являє собою сітківку. Сітківка складається зі світлочутливих (сенсорних) нервових та опірних клітин (мал. 64).
Мал.10. Схематичний розріз через сітківку ока (за Поляковим): 1 - пігментний шар; 2 - шар фоторецепторів (а - палички, б - колбочки; З — зовнішній сітчатий шар; 4 - шар біполярних клітин - внутрішній ядерний шар; 5 - внутрішній сітчатий шар; 6-шар гангліозний клітин; 7-шар нервових волокон, що йдуть у зоровий нерв. До пігментного шару прилягає шар фоторецепторів, зовнішні членики яких мають вигляд паличок і колбочок. Ядра світлочутливих клітин утворюють зовнішній клітинний шар, а далі йде зовнішній сітчастий шар, де кінцеві розгалуження фоторецепторів контактують з дендритами біполярних клітин. Відростки останніх контактують з дендритами гангліозних клітин у внутрішньому сітчастому шарі. Гангліозні клітини дають довгі нервові волокна (аксони), що утворюють внутрішній шар сітківки і далі складові зорового нерва. Подальший розвиток парних органів зору зводиться до вдосконалення світлозаломлюючих середовищ, розвитку скелетної оболонки та набуття мускулатури, яка служить для приведення в рух очного яблука. У ряду хребетних акомодація поступово вдосконалюється, спостерігається набуття рухомих повік, слізних залоз та інших частин ока. Прогресивна також поява об'ємного бачення (стереоскопія) у деяких ссавців.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 703; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.42.233 (0.014 с.) |