
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Органи дихання первинноводних організмів. ПоходженняСодержание книги
Поиск на нашем сайте
У хордових для дихання пристосований передній відділ кишечника, продірявлений більш-менш численними отворами чи щілинами, що ведуть назовні або безпосередньо, або через особливу навколозяброву порожнину. Дихальна течія води підтримується в нижчих формах рухами миготливих війок, а у вищих (хребетних) - мускульними дихальними рухами. У хребетних, як і в нижчих хордових, органи водного дихання розвиваються в передньому відділі кишечника, ембріонально - в ділянці глотки. Тут з'являються парні кишенеподібні вирости ектодермальної стінки, які ростуть у напрямі до покривів, звідки їм назустріч розвиваються відповідні ектодермальні випини. Внутрішні вирости сходяться з зовнішніми випинами, їх стінки на місці стикання зростаються, а потім утворюють прорив.'Отже, розвивається парний ряд вісцеральних мішків, що відкриваються назовні за допомогою щілин. Вісцеральні щілини (або, принаймні, їх кишенеподібні закладки) розвиваються спочатку в усіх хребетних, але в наземних вони потім заростають і зникають. У нижчих хребетних у стінках цих вісцеральних мішків розвиваються зябра. У круглоротих зяброві мішки здуваються й набирають вигляду сочевицеподібних порожнин. Мішок внутрішнім вузьким каналом відкривається в порожнину глотки, а зовнішнім - на бічну поверхню тіла тварин. На ентодермальній стінці цих мішків слизова оболонка утворює меридіональні зяброві складки (мал. 1).
Мал. 1. Органи дихання морської міноги: 1 - зяброві мішки; 2 -рогові зуби; З - загальний зябровий прохід; 4 - внутрішні та 5 - зовнішні зяброві проходи; 6 -рот; 7 - зовнішні зяброві отвори; 8 — перегородки між зябровими мішками.
Дихання здійснюється шляхом ритмічних стискань і розслаблень м'язової стінки зябрової решітки, яка при вдиху розправляється і вода заходить у глотку і в мішки знову. У риб зяброві мішки мають вигляд щілиноподібних просторів, що лежать між зябровими перегородками (мал. 2, а, б).
2 - клапани міжзябрових перегородок; 3 - зяброві пелюстки; 4 -ротова порожнина; 5 - зовнішні зяброві порожнини; 6 - зяброва кришка.
В останніх ембріонально є ділянка порожнини тіла у вигляді каналу, і розвивається свій скелет у вигляді вісцеральних дуг, які лежать у внутрішній частині перегородки. На вісцеральних дугах розвиваються складки слизової оболонки - зяброві пелюстки; їх сукупність складає зябра. У хрящових риб до під'язикової та зябрових дуг прикріпляються міжзяброві перегородки, на бічних поверхнях яких розміщені зяброві пелюстки ектодермального походження. В основі кожної міжзябрової перегородки проходить приносна зяброва артерія (має венозну кров), яка розпадається на капіляри в зябрових пелюстках обох півзябер. Капіляри об'єднуються в парні виносні зяброві артерії (артеріальна кров), які над зябровою щілиною об'єднуються в загальну виносну артерію, по останній тече кров, збагачена киснем. У деяких акул кожна перегородка продовжується ззовні в шкірну складку, що прикриває зяброву щілину, яка йде ззаду (мал.65, б). Початкова функція цих складок - захист зябер від попадання сторонніх тіл ззовні. Однак вони також беруть участь у дихальних рухах як розріджуючий насос, і при прогресивному їх розвитку ця роль стає дедалі активнішою. Зяброві перегородки й складки підтримуються хрящовими променями, що відходять від вісцеральних дуг. Зябровий апарат вищих риб захищений ззовні зябровою кришкою (operculum), яка підтримується власним кістковим скелетом. У зв'язку з цим змінюється й будова зябер, розміщених тільки на I-IV скостенілих зябрових дугах. Міжзяброві перегородки редукуються і практично зяброві пелюстки обох півзябр прикріплюються до дуги і з сидячих перетворюються в гребенясті зябра. В основі зябрових пелюсток лежить приносна зяброва артерія, відгалуження якої розпадаються на капіляри, що йдуть в кожну зяброву пелюстку; вони зливаються у виносну зяброву артерію, яка несе окислену кров і впадає в корінь аорти. Кінці півзябр послідовних зябрових дуг при житті тримаються зімкнутими, і вода проціджується при дихальних рухах через складну систему щілин, що лежать між вторинними пелюсточками. Дихальна поверхня зябер досягає, таким чином, значної величини і складає 1-3 см2 на 1 г маси риби. Газообмін між водою і кров'ю в зябрових пелюстках здійснюється за допомогою численних капілярів, кров у яких тече назустріч потоку води, що омиває зяброві пелюстки. Це помітно підсилює газообмін. Завдяки такій системі риби здатні засвоювати 46-82 % розчиненого у воді кисню. Одночасно через зябра виділяється вуглекислий газ і деякі продукти метаболізму (сечовина, аміак). Зябра беруть участь у сольовому та водному обміні. В ембріонів кисень засвоюється добре розвиненою сіткою кровоносних судин жовточного мішка та плавцевої складки. У личинок поряд з цим на плавцях, голові, боках тіла функціонує сітка капілярів, а в деяких видів (дводишні риби, багатопері) розвиваються зовнішні зябра - вирости шкіри, густо пронизані кровоносними судинами. Поступово основна функція дихання переходить до внутрішніх зябер. У наземних хребетних закладається, як і в риб, до шести вісцеральних мішків. З першого мішка, що відповідає бризкальцю, розвивається порожнина середнього вуха, наступні зяброві мішки прориваються ще звичайно назовні, але потім зникають, даючи лише початок деяким похідним їх епітелію (наприклад, зачатку щитовидної залози). Тільки в амфібій розвиваються справжні зябра як личинкові органи дихання. У хвостатих амфібій - це зовнішні зябра, а в безхвостих потім розвиваються і внутрішні ектодермальні зябра. Закладається й зяброва кришка у вигляді шкірної складки, що прикриває основи зовнішніх зябер. У личинок безхвостих амфібій зяброва складка розростається, прикриваючи весь зябровий апарат, а оперкулярна щілина зменшується до ступеня невеликого отвору, який звичайно зростається з отвором другого боку й розміщається асиметрично. У дорослих амфібій зяброві щілини заростають (за винятком неотенічних форм).
Додаткові органи газообміну
У кісткових риб, які живуть у водоймах, де спостерігаєть-ся постійний або періодичний дефіцит кисню, наявні додаткові органи дихання. Перш за все таким органом служить шкіра. У риб, пристосованих жити в умовах значного дефіциту кисню, частка шкірного дихання в загальному газообміні досягає 17-22 %, а в окремих видів 42-80 %. Такими є в'юни, карасі, вугрі, соми тощо. При цьому риби, в яких шкіра має найбільше значення в диханні, позбавлені луски, або ж вона дрібна й не утворює суцільного покриву. У різних систематичних групах риб розвиваються пристосування до використання атмосферного кисню. При дефіциті кисню у воді багато коропових захоплюють повітря ротом. При цьому вода в ротовій порожнині аерується, а кисень повітряних пухирців поглинається зябрами й слизовою оболонкою ротової порожнини. У деяких тропічних костистих риб є лабіринтовий апарат -розширена кишенеподібна ділянка зябрової порожнини, утворена системою складок слизової оболонки, густо пронизаних капілярами. Лабіринтові риби дихають атмосферним киснем і можуть упродовж кількох днів жити на суші (анабас, макроподи, гурамі та ін.). Для деяких коропових (пічкурів, в'юнів) і сомових характерне кишкове дихання. Для цього служать спеціалізовані ділянки кишки, позбавлені травних залоз. Риба захоплює повітря ротом, пропускає його через травний тракт завдяки перистальтиці; при цьому засвоюється до 50 % кисню й виділяється вуглекислий газ. Нарешті, як додатковий орган дихання служить іноді плавальний міхур. У променеперих риб він розвивається як порожнистий тонкостінний виріст спинної поверхні передньої частини стравоходу. При цьому в так званих відкри-томіхурних риб (кісткові й хрящові ганоїди, оселедцеподібні, коропоподібні) плавальний міхур зберігає зв'язок зі стравоходом, а в закритоміхурних (окунеподібні та деякі інші) невдовзі після вилуплення личинки міхур цілком відокремлюється від стравоходу. У дводишних і китичноперих риб роль плавального міхура виконують легені, що утворюються як парний виріст черевної стінки стравоходу; тільки в рогозуба він вторинно став непарним. Ці легені забезпечуються кров'ю кровоносними судинами, що відходять від п'ятої виносної зябрової артерії і впродовж усього життя зберігають зв'зок із стравоходом. Стінки легень комірчасті, що збільшує поверхню окислення. Отже, досконалий механізм зябрового дихання, розвиток різноманітних додаткових органів дихання, що дозволяють використовувати атмосферний кисень, забезпечили можливість широкого розселення кісткових риб по водоймах з різним вмістом кисню, та газовим режимом. Серед наземних хребетних додаткові органи дихання характерні для земноводних. У дорослих форм це шкіра й частково слизова оболонка ротової порожнини. При цьому у видів, що живуть у більш вологих місцях, шляхом шкірного дихання поглинається до 50 % кисню й виділяється до 86 % вуглекислого газу; у видів, що живуть у більш сухих умовах, ці показники складають відповідно 34 і 74 %. У личинок амфібій у газообміні беруть участь шкіра, зовнішні та внутріщні зябра, плавцеві складки.
Органи дихання тетрапод. Походження Легені (pulmones) - це органи дихання, характерні для деяких риб (дводишних, китичноперих, багатоперих), наземних хребетних і людини. У нижчих хребетних (анамній) легені закладаються у вигляді парного випину на черевній стінці глотки як гомолог останньої пари зябрових мішків. З подальшим розвитком ці зачатки ростуть назад, видовжуючись у мішкоподібні утвори. В одночас відокремлюється ділянка глотки, що сполучає обидва зачатки. Вона дає початок непарному дихальному шляху (гортані й трахеї), а
Мал.3. Закладка легень у амфібій (за Гетте): 1 -5 - вісцеральні мішки; 7 — закладка легень; 8 - закладка зовнішньої зябри; 9 - порожнина глотки; 10 - порожнина тіла; 11 -ротова ямка. У вищих наземних хребетних (амніот) дихальний шлях звичайно закладається раніше ніж легеневі мішки, і розвиток починається з відокремлення непарного жолоба на черевній стінці глотки ззаду закдадок зябрових мішків. На цьому жолобі закладаються потім самі легені у вигляді парного випину. Крім того, і в самій легені найраніше закладаються дихальні шляхи - бронхи та їх розгалуження. Тому процес розвитку легень являє собою картину поступового утворення нових гілок, що мають кінцеві здуття. Легені забезпечуються кров'ю легеневою артерією, гомологічною IV зябровій артерії риб. Схожість легень нижчих наземних хребетних з плавальним міхуром деяких риб уже давно навела дослідників на думку про гомологію цих утворів. Однак цей погляд у такій загальній формі натрапляє на чималі труднощі. Найбільш важливий аргумент, що легені виникли раніше, ніж плавальний міхур. Ми бачили, що плавальний міхур більшості риб є непарним органом, який розвивається в спинному мезентерії. Він постачається артеріальною кров'ю кишкових судин (а. соеііаса) і віддає венозну кров почасти в кардинальні, почасти у ворітну вену печінки. Ці факти, безперечно, свідчать проти зазначеної гіпотези. Однак у деяких риб плавальний міхур парний і постачається, як і легені наземних хребетних, кров'ю від четвертої пари зябрових артерій та віддає її прямо в серце. Цілком зрозуміло, що ми маємо тут справу з утворами того ж роду, що й легені. Спираючись на факт закладання легень в амфібій у вигляді парних мішкоподібних випинів поззаду від останьої пари зябрових мішків, Гетте висловив припущення, що легені являють собою результат перетворення пари зябрових мішків. У даний час загальновизнаною є теорія, що доводить походження легень наземних хребетних від спеціалізованих легень давніх кистеперих риб. Як було вже сказано, легені дводишних риб є лише додатковим органом, бо нормально ці риби дихають зябрами. Тільки в період засухи, коли тварина заривається в мул (протоптеруси), легеневе дихання набуває важливого значення. Легені китичноперих ганоїдів (поліптерусів) мають гладенькі стінки й відіграють малопомітну роль навіть як додаткові органи дихання. Те саме стосується й деяких амфібій. У постійнозябрових земноводних (Nesturus, Proteus) легені мають вигляд видовжених гладкостінних мішків, як і в поліптерусів, і при тому також асиметрично розвинені. І тут вони переважно відіграють роль гідростатичного апарату. У багатьох інших амфібій легеневе дихання ще надто незначне(15 %). Переважає дифузне шкірне дихання, а в безлегеневих саламандр легені відсутні. Однак у більшості хвостатих амфібій внутрішня поверхня легень збільшена внаслідок утворення сітки перекладин, що обмежовують систему комірок (мал.4).
Мал. 4. Схема будови легень у хребетних (за Відергеймом): І-ІІ - хвостаті земноводні; НІ - безхвості земноводні; IV - лускаті плазуни; V- крокодил і черепаха; VI-птахи; VII-ссавці. 1 - бронхи; 2, 3 - черевні та спинні гілки бронха; 4 - легеневі альвеоли; 5 - легеневі трубки; б — з'єднання легень з повітряними мішками. У безхвостих амфібій легені мають вигляд мішків зі складною системою комірок, бо всередині окремих більших комірок розвивається система вторинних перекладин, які обмежовують систему дрібніших комірок. Отже, стінка легеневого мішка може набрати майже губчастого вигляду. Центральна частина легеневого мішка має характер дихального шляху. У земноводних повітря нагнітається в легені з ротової порожнини ритмічними скороченнями її дна при закритій гортанній щілині та закритих ніздрях, другий акт - розслаблення м'яза й відкривання гортанної щілини, при цьому повітря із легень поступає в рот, змішується зі свіжим. Далі м'яз скорочується і заганяє змішане повітря в легені. Ще один акт - скорочується м'яз, відкривається щілина й ніздрі, відбувається видих. В амніот з розвитком значного рогового покриву шкіри легені набувають значення єдиного органа дихання й відповідно диференційовані значно вище. Стінки легень стають губчастими, а центральна порожнина диференціюється на систему внутрішньолегеневих дихальних шляхів. У нижчих плазунів (Hatteria) стінки легень комірчасті й дуже схожі на легені амфібій, хоч верхівка вже трохи виступає вперед. У деяких ящірок, черепах, крокодилів легені мають губчасту будову, тобто перекладини, розростаючись, ділять порожнину легені на камери й комірки, всередині яких розвиваються вторинні перекладини, що обмежовують систему дрібних комірок, збільшуючи дихальну поверхню легень. Вільним залишається центральний прохід – внутрішньолегеневий бронх (мал. 68). У деяких плазунів (хамелеонів, геконів, деяких ящірок і змій) легені продовжуються в тонкостінні додатки – легеневі мішки. Ці резервуари повітря забезпечують ефект шипіння, полегшують газообмін при тривалому проходженні їжі по стравоходу та при пірнанні. Вентиляція легень у більшості плазунів забезпечується роботою грудної клітки з допомогою міжреберної та черевної мускулатури. В акті дихання, особливо в черепах, беруть участь плечові й тазові м'язи. У черепах зберігається й ротоглотковий механізм нагнітання повітря, характерний для земноводних. У вищих наземних хребетних процес відокремлення дихальних шляхів всередині самої легені йде значно далі, в результаті чого розвивається складна система бронхів різного порядку. Легені птахів прирощені до спинного боку сторони грудної клітки. Це малорозтяжні губчасті органи. Бронхи входять у кожну легеню з черевного боку й продовжуються у вигляді головного внутрішньолегеневого бронха до заднього кінця легені, а поза легенею переходять у порожнину тонкостінного черевного повітряного мішка. Від головного бронха відходять вторинні бронхи. Як правило чотири з них пронизують усю легеню й продовжуються за її межами у вигляді повітряних мішків (мал. 68).
Крім того, певна кількість вторинних бронхів (10-14) закінчуються в легенях сліпо. Між вторинними бронхами розміщуються з'єднуючі їх численні анастомози - парабронхи, від яких відходить безліч радіально розміщених каналів з комірчастими стінками, густо обплетеними капілярами. Це бронхіоли, де відбувається газообмін. Загальна дихальна поверхня легень птахів значно перевищує дихальну поверхню легень плазунів і може бути зіставлена з такою у ссавців. З легенями птахів, як було сказано, зв'язані повітряні мішки - прозорі еластичні тонкостінні вирости слизової оболонки вторинних бронхів. їх об'єм в декілька разів перевищує об'єм легенів. Розміщені вони між внутрішніми органами, а їх відростки проникають під шкіру та в порожнини трубчастих кісток. Повітряні мішки забезпечують подвійну вентиляцію легень з газообміном при вдиху та видиху. У зв'язку з таким досконалим диханням і оксидація відбувається енергійніше, і звільняється значна кількість тепла. Птахи мають сталу, при тому дуже високу (39-43°С) температуру тіла. Повітряні мішки здійснюють інтенсивну тепловіддачу під час польоту, запобігаючи перегріву організму (Шмідт-Нієльсен, 1982). У легенях ссавців внутрішньолегеневе бронхіальне дерево галузиться на велику кількість вторинних бронхів, а останні -на бронхи третього й четвертого порядків, що закінчуються тонкостінними трубчастими бронхіолами, які відкриваються в альвеолярні ходи з кінцевими здуттями альвеолярними міхурцями та альвеолами (мал.68). Газообмін відбувається в легеневих альвеолах, які вистелені плоским дихальним епітелієм і мають багату сітку капілярів. Така структура легень збільшує їх дихальну поверхню, яка в 50-100 разів перевищує поверхню тіла. Механізм дихання у ссавців подвійний. Акт дихання здійснюється за допомогою періодичного розширення та стиснення грудної порожнини внаслідок: 1) зміни положення ребер за допомогою реберної мускулатури; 2) скорочення й розслаблення діафрагми, яка має форму купола. При видиханні певну роль відіграє також еластичність легеневих міхурців і наявність гладеньких мускульних волокон, які особливо добре розвинені в китоподібних. Отже, легені хребетних, що виникли як додаткові органи дихання в риб у вигляді мішкоподібних виростів останньої пари зябрових мішків, зберегли це значення у дводишних та китичноперих риб. У наземних хребетних вони поступово набули значення єдиного органу дихання. В амфібій легені ще не відіграють великої ролі й мало чим відрізняються від легень риб. Прогресивна еволюція легень проявляється у відокремленні більш чи менш складних дихальних шляхів і в збільшенні дихальної поверхні внаслідок розвитку дедалі складнішої сітки перекладин, які надають стінкам легень губчастої будови. Більші перекладини розбивають легені на окремі ділянки, всередині яких відокремлюються центральний дихальний шлях і периферична комірчаста частина, в якій у свою чергу відбувається така ж диференціація. Врешті-решт, розвивається складна система розгалужень внутрішньолегеневих бронхів, які закінчуються кінцевими міхурцями з комірчастими стінками (з легеневими альвеолами). Поступово вдосконалюється механізм дихання - від нагнітального в амфібій і деяких плазунів до міжреберно-діафрагмального у ссавців. У результаті цього вдосконалювалась вентиляція легень й оксидація відбувалась все енергійніше.
Дихальні шляхи
Дихальні шляхи являють собою по суті більш чи менш довгу трубку, за допомогою якої легені сполучаються з порожниною глотки. На відміну від дихальних частин самих легень, дихальні шляхи вислані циліндричним миготливим епітелієм. Сукупність хрящів, м'язів і зв'язок, що утворюють передній відділ дихального горла, має назву гортані (larynx). Порожнина гортані поділена на передню частину, яка через дихальну щілину з'єднується з порожниною глотки, і задню, яка переходить в порожнину трахеї. Органами опори для початкового відділу є гортанні хрящі, що являють собою продукт перетворення останніх зябрових дуг. Сама ж гортань - похідна глотки. У більшості хвостатих та в усіх безхвостих земноводних розрізняють черпалоподібні (cartilago arytaenoideae) і перснеподібні (cart, cricoidea) хрящі. У безхвостих земноводних гортань зливається з трахеєю й утворює короткий гортанно-трахеальний мішок.
Уздовж внутрішніх країв черпалоподібних хрящів є складки слизової оболонки, розділені голосовою щілиною. Знизу до перснеподібного хряща приростає півкільце, утворене хрящами трахеї. Увесь цей утвір називається перснеподібно-трахеальним хрящем. Гортань плазунів і птахів має ті ж самі хрящі, але складнішої будови. Значно краще розвинена гортань у ссавців, у яких вона є й голосовим апаратом. Крім черпалоподібних та перснеподібних хрящів є ще щитоподібний (cart. thyreoidea), розміщений з черевного боку, спереду від кільцеподібного хряща, і надгортанник (epiglottis), який сидить на ньому ще далі спереду в непарній складці слизової оболонки. Щитоподібний хрящ у клоакових чітко виявляє свій склад із трьох пар хрящів, які, безперечно, є першими трьома парами зябрових дуг, і утворює одне ціле з під'язиковим апаратом. У живородних ссавців також неважко встановити походження щитоподібного хряща на основі його розвитку - тут до його складу входять лише дві пари зябрових дуг, друга й третя, тим часом як передня їх пара прилягає до щитоподібного хряща спереду, не втрачаючи своєї самостійності. Це - задні ріжки під'язикового апарату. Походження надгортанника не зв'язане з вісцеральним апаратом. Він розвивається в складці слизової оболонки перед гортанною щілиною. Голосові зв'язки натягнені між черпакуватими та щитоподібними хрящами. Будова гортані залежить від особливостей живлення й дихання тварин. Так, у китоподібних і новонароджених сумчастих ссавців дуже виражені черпакуваті хрящі й надгортанник утворюють трубку, яка вдається позаду м'якого піднебіння в хоани. Така будова гортані дозволяє сумчастим одночасно дихати і ссати молоко; у китоподібних вона запобігає потраплянню в гортань води при ковтанні їжі. У деяких хвостатих земноводних є вже трахея (trachaea) у вигляді досить довгої трубки. У земноводних трахея не розділяється на бронхи й легені починаються від трахеї. У безхвостих амфібій трахея відсутня, а легені починаються. від гортані. Стінки трахеї зміцнюються нерівномірно розкиданими хрящиками, що іноді сполучаються по боках між собою. Ці хрящики виявляють певний зв'язок з гортанними хрящами, тому їм приписують таке ж вісцеральне походження, як і останнім. У плазунів, птахів і ссавців трахея добре розвинена й поділяється на бронхи. Довжина трахеї прямо пропорційна довжині шиї та обернено пропорційна довжині бронхів. У плазунів скелет трахеї складається з повних або неповних хрящових кілець. У птахів трахейні кільця звичайно повні й іноді костеніють. Щитоподібний хрящ у клоакових чітко виявляє свій склад із трьох пар хрящів, які безперечно становлять три пари зябрових дуг й утворює одне ціле з під'язиковим апаратом. У живородних ссавців також не важко встановити походження щитоподібного хряща на основі історії його розвитку - тут до його складу входять лише дві пари зябрових дуг, друга й третя, тим часом як передня їх пара прилягає до щитоподібного хряща спереду, не втрачаючи своєї самостійності. Це - задні ріжки під'язикового апарату. Походження надгортанника не зв'язане з вісцеральним апаратом. Він розвивається в складі слизової оболонки спереду від гортанної щілини. Нерідко трахея видовжується ще більше, ніж шия, й утворює складні згини, що розміщуються під шкірою або всередині гребеня груднини. Така довга трахея відіграє роль резонатора. Голосовим апаратом птахів є нижня гортань (syrinx), що лежить на межі між трахеєю та бронхами. Джерелом звукових коливань є різно збудовані перетинки, що розміщуються почасти в ділянці бронхів, почасти в трахеї. Крім того, часто спостерігається складка з кістковим або хрящовим скелетом - козелок (pessulus), яка вдається в порожнину барабана від місця сходження внутрішніх стінок бронхів. Нижня гортань має свою більш-менш складно диференційовану мускулатуру, скороченням різних м'язів якої змінюється натяг окремих перетинок і регулюється висота звука. Ця мускулатура розвивається (на відміну від вісцеральної мускулатури гортані) через відокремлення від під'язикової мускулатури. Трахея ссавців має значну довжину, підтримується хрящовими кільцями (звичайно неповними) й поділяється на два виразних бронхи, що підтримуються такими ж кільцями. Трахея вистелена зсередини слизовою оболонкою, під якою міститься багато слизових залоз, і вкрита миготливим епітелієм, що перешкоджає проникненню в легені пилу й сторонніх часточок. Ззовні трахея вкрита сполучнотканинною, або серозною (у грудній порожнині) оболонкою. У загальному плані еволюція дихальної системи спрямована на: збільшення поверхні дотику з навколишнім середовищем і підвищення проникності стінок; диференціацію дихальних шляхів, що зумовлює більш досконале проходження та газообмін повітря; оптимальний контакт атмосферного повітря з кров'ю через стінки альвеол.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 792; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.186.26 (0.013 с.) |


 Мал. 2. Схематичні горизонтальні розрізи через голову костистої риби (А) та акули (Б). Зліва - положення клапана при вдиху, справа - при видиху (за Воскобойніковим): 1 - гіоїднип клапан;
Мал. 2. Схематичні горизонтальні розрізи через голову костистої риби (А) та акули (Б). Зліва - положення клапана при вдиху, справа - при видиху (за Воскобойніковим): 1 - гіоїднип клапан;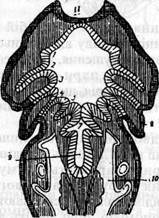 парні мішки вростають у черевну порожнину, покриваючись при цьому плеврою, і перетворюються в органи більш чи менш складної будови. Завдяки розвитку дедалі складнішої сітки перекладин внутрішня дихальна поверхня легеневих мішків поступово збільшується. (мал. 3).
парні мішки вростають у черевну порожнину, покриваючись при цьому плеврою, і перетворюються в органи більш чи менш складної будови. Завдяки розвитку дедалі складнішої сітки перекладин внутрішня дихальна поверхня легеневих мішків поступово збільшується. (мал. 3).



