Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекція 4. М’язова система хордових тварин (2 год.)Содержание книги
Поиск на нашем сайте
Логіка викладу. 1. Соматична мускулатура хордових тварин. ЇЇ розташування та функції. 2. Вісцеральна мускулатура хордових тварин. ЇЇ розташування та функції. 3. Диференціювання мускулатури у вищих хордових тварин. Рекомендована література: 1, 2, 3, 4, 5.
ФУНКЦІЯ: М'язова система здійснює процеси руху всередині організму (робота серця, судин, шлунка, кишечника тощо), переміщає організм або його частини в просторі, забезпечує опорну функцію, здійснює механічну роботу, генерує і проводить нервове збудження (провідна система серця), виробляє електричну енергію в деяких риб. Нагадаємо, що у всіх представників Типу Хордові скелетна мускулатура представлена посмугованими м'язами. У хордових виявляється поділ мускулатури на соматичну, яка зумовлює рух тіла, і вісцеральну, що зумовлює рухи внутрішніх органів. Такий поділ можна провести у всіх тварин, хоча звичайно між цими групами м'язів немає помітних відмінностей в їх походженні та іннервації. Але в хордових тварин відмінність між соматичною й вісцеральною мускулатурою виявляється значно глибшою. У ланцетника соматична мускулатура розвивається за рахунок внутрішньої стінки сомітів. У дорослої тварини вона має вигляд поздовжнього бічного м'яза, поділеного драглистими перегородками на ряд сегментів (міомерів), що відповідають ембріональним міотомам. Гладенька мускулатура судин та кишечника розвивається за рахунок внутрішнього листка несегментованої мезодерми. У хребетних чітко виражений поділ на соматичну (парієтальну) і вісцеральну мускулатуру, які відрізняються як своїм розвитком, так і іннервацією. Кожне мускульне волокно зв'язане з певним нервовим волокном й утворює разом з ним одну фізіологічну одиницю. Отже, цілий м'яз нерозривно зв'язаний з певним нервом. Парієтальна м'язова система
Парієтальна, скелетна, або соматична мускулатура розвивається за рахунок внутрішнього листка сегментованої частини мезодерми й іннервується спинномозковими нервами. Уся вона складається з поперечносмугастих м'язів, що забезпечують взаємодію тварини з довкіллям. Соматична мускулатура хребетних включає м'язи тулуба, кінцівок та їх поясів, м'язи очного яблука та під'язикової дуги, діафрагми. Мускулатура тулуба в нижчих хребетних складається з ряду міомерів правого й лівого боків тіла, які разом утворюють бічні м'язи. Бічний м'яз кожного боку поділений горизональною сполучнотканинною перегородкою на спинний і черевний м'язи; відповідно кожен міомер поділений на спинний і черевний відділи, надосьовий (епіаксіальний) і підосьовий гіпаксіальний) тяжі. Кожен міомер іннервується руховою гілкою спинномозкового нерва: спинний відділ - спинною гілкою, черевний - черевною. Міомери дорослої тварини утворюють складну систему конусів, що вкладаються один в одного і мають у поперечному розрізі вигляд концентричних кілець. Початковий поздовжній напрям волокон вже у риб починає змінюватись у ділянці черевного відділу мускулатури. У бічних стінках черева зберігається поздовжній напрям волокон. Тобто, починається вже деяка диференціація всередині самих міомерів. Це приводить до відокремлення різних шарів черевної мускулатури. У наземних хребетних прикріплення міосепт точно пристосовується до окремих сегментів скелета, що пов'язане з розвитком рухомого зчленування між хребцями. В амфібій спинна мускулатура складається ще з ряду незалежних міомерів, поділених, як і в риб, міосептами на спинний і черевний відділи. В амніот горизонтальні септи зникають і межа між черевним і спинним відділами також зникає. Починаючи з плазунів, міомери почасти розбиваються па окремі мускульні пучки, що зберігають метамерний розподіл і зв 'язують між собою окремі частини сусідніх хребців, почасти сполучаються між собою для утворення бічних м 'язів. Саме так розвиваються міжостисті м'язи (m.interspinalis), що сполучають верхні остисті відростки; поперечні м'язи (т. transversus), що сполучають між собою поперечні відростки. Від поперечних відростків до ребер йдуть поперечнореберні м'язи (m.transversocostales), а від передніх хребців до голови - прямі й косі (m. recti et obliqui capitis). Вздовж усієї спини в кутку між остистими и поперечними відростками лежить довгий м'яз спини (m. longissimus dorsi), передня частиня якого може відокремитися у вигляді довгого м'яза голови (т. longissimus capitis). Над ребрами розвинена система м'язів (m. ileocostales). Ззаду тазового пояса диференціюється особлива хвостова мускулатура (почасти за рахунок черевної мускулатури), залишки якої спостерігаються й у безхвостих форм, включаючи людину. У результаті цього диференціювання від початкової метамерії спинної мускулатури у всіх вищих хребетних зберігаються тільки сліди у вигляді метамерного розподілу окремих м'язів, що зв'язують між собою послідовні хребці. Значення спинної мускулатури, яка в риб є найважливішою частиною локомоторного апарату, в наземних хребетних значно менше. Замість цього досить сильно розвивається мускулатура кінцівок. У нижчих форм наземних хребетних спостерігається чітке диференціювання черевної мускулатури на відділи й шари, що відрізняються напрямом волокон (мал.35).
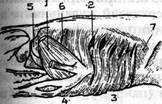
Мал. 35. Поверхнева черевна мускулатура тритона (за Маурером). 1-підзябровий м'яз; 2-міосепта; 3-зовнішній косий м 'яз; 4-внутрішній прямий м 'яз; 5-загальпий прямий м 'яз черева.
Посередині черева відокремлюється поздовжня мускульна смуга - прямий м'яз черева (m.rectus abdominis), no середній лінії якого є сполучнотканинна перегородка, яка відділяє мускулатуру правого й лівого боку. У бічних стінках черева напрям волокон змінюється, причому по-різному на різній глибині: у поверхневих шарах волокна напрямлені косо назад до черева, у глибших вони йдуть косо назад до спини, а в найглибших набирають поперечного напряму. У нижчих земноводних ці три шари ще зв'язані один з одним і диференціювання виражене лише в різному напрямі волокон. У більш високоорганізованих форм ці шари цілком відокремлюються один від одного й відповідно утворюються зовнішній косий м'яз черева (m. obliquus externus), внутрішній косий м'яз черева (m. obliquus interims) та поперечний м'яз (m. transversus abdominis). Поздовжні волокна прямого м'яза, що лежать по середній лінії, також цілком відокремлюються від сусідніх м'язів. У вищих земноводних, як і в інших наземних хребетних, міосепти зникають і мускульні волокна в косих та поперечному м'язах йдуть без перерви на всьому їх протязі. Сліди колишньої сегментації зберігаються лише в прямому м'язі живота у вигляді сухожильних тяжів, що йдуть поперек м'яза (у людини їх 3-4). З розвитком ребер в амніот черевні м'язи поділяються в грудній ділянці на окремі частини, що лежать між ребрами. Це міжреберні м'язи, зовнішній і внутрішній, що є безпосереднім продовженням косих м'язів черева й відіграють велику роль при дихальних рухах (мал.36).
Мал. 36. Черевна мускулатура гатері і (за Маурером). М'язи: міжзябровий; 2-поверхневий зовнішній косий; 3-грудний; 4 - прямий м 'яз черева; 5 - черевне ребро; 6 - груднина.
У поперековій ділянці їм відповідає квадратний поперековий м'яз, що продовжується й під міжреберними м'язами та примикає збоку до поперечного. У ссавців зовнішній і внутрішній міжреберні м'язи дають початок ще новим продуктам диференціювання - заднім зубчастим м'язам, верхньому й нижньому (mm. serrati postici superior et inferior), які спеціалізуються як дихальні м'язи, що служать для розширення грудної клітки. У сумчастих з черевного боку від прямого м'яза черева відокремлюється ще особливий утвір, який відходить від сумчастих кісток і доходить спереду до груднини. Це пірамідальний м'яз (m. pyramidalis), який є й у нижчих плацентарних ссавців, а в інших зберігається в вигляді рудимента, зв'язаного з лобковими кістками. Усе це диференціювання обумовлене зміною функції черевної мускулатури. Якщо в риб вона, як і спинна мускулатура, виконує локомоторну функцію, то зі зміною способу пересування й набуттям легеневого дихання - служить для стискання черевної порожнини й, зокрема, для виконання дихальних рухів. Мускулатура непарних плавців розвивається у вигляді метамерного ряду епітеліальних виростів спинних або черевних кінців міотомів. Ці мускульні пупки вростають в основу закладки плавця, втрачають зв'язок з міотомами й перетворюються в ряд синцитіїв, у яких потім утворюються мускульні волокна. Таким чином, відповідно до кожного променя внутрішнього скелета розвивається з кожного боку по одному первинному променевому м'язу. М'язи плавців прикріплюються завжди до променів шкірного скелета, іннервуються такою кількістю спинномозкових нервів, скільки міотомів брало участь у їх розвитку. Мускулатура парних плавців закладається також у вигляді ряду мускульних пупків, що виростають від нижніх кінців міотомів. Кожний мускульний пупок поділяється на верхній і нижній вторинні пупки, які вростають в основу закладки плавця зі спинного й черевного боку й перетворюються в такі ж первинні променеві м'язи, як і в непарних плавцях. Променеві м'язи тісно зближаються між собою й утворюють суцільний мускульний шар на спинному й черевному боці плавця. Мускулатура кінцівок наземних хребетних розвивається також з ряду мускульних пупків, які швидко втрачають свої контури й розпливаються в масу мезепхіматозних клітин, що утворюють суцільний шар на тильній і долонній сторонах зачатка кінцівки. У межах цих мускульних зачатків відбувається потім диференціювання, причому мускульні зачатки передньої кінцівки утворюють проксимальні вирости, що розростаються в ділянці плечового пояса на спину й на груди, утворюючи м'язи плечового пояса. М'язи, що розвиваються з цих первинних мускульних зачатків, гомологічні м'язам парних плавців і становлять первинну мускулатуру кінцівок. Із спинного зачатка мускулатури, гомологічного відвідному м'язові плавця, розвивається група розгиначів п'ятипалої кінцівки, а з черевного зачатка, гомологічного привідному м 'язові плавця, розвивається система згиначів. Крім цієї первинної мускулатури як продукт дальшого диференціювання міотомів грудної ділянки, розвивається ще особлива вторинна мускулатура плечового пояса, яка служить для його пересування. Із спинного мускульного зачатка розвиваються в передній кінцівці такі первинні м'язи: дельтоподібний (т. deltoideus) спереду, спинний м'яз (т. dorsalis scapulae) над лопаткою, підлопатковий (m. subscapularis) і широкий м'яз спини (т. latissimus dorsi) ззаду; у межах вільної кінцівки - триголовий м'яз (m. anconeus), що йде від лопатки, коракоїда та плеча до ліктьового відростка, та всі розгиначі кінцівки, включаючи розгиначі пальців (m. extensores). З черевного мускульного зачатка розвиваються в передній кінцівці такі м'язи (мал.37): черевні м'язи плечового пояса (т. coraco-brachialis) спереду та грудний м'яз (m. pectoralis) ззаду; в межах вільної кінцівки - коракоїдно-променевий м'яз (т. coraco-radialis) та всі згиначі (m. flexores) кінцівки.
Мал. 37. Поверхневі м 'язи передньої частини саламандри (за Фюрбе-ріигером).1 — три -чотириголовий м'яз; 2 - трапецісвидний; З -м'яз, що опускає щелепу; 4 - спинний; 5 -зовнішній косий; 6 - широкий м 'яз спини; 7-м 'яз, що піднімає лопатку; 8 -жувальний; 9 - міжщелепний; 10 - підборідно-під 'язиковий; II - грудний; 12 -прокоракоїдно-плечовий; ІЗ - прямий черевний; 14 — падкоракоїдний. Вторинна мускулатура плечового пояса складається із значного м'яза, що розвивається спереду пояса - він піднімає лопатку (m. levator scapulae), і передніх зубчастих м'язів, що розвиваються позаду (m. serrati antici superficialis et profundus). М'яз, що піднімає лопатку, є в акул, але відсутній у вищих риб, у яких пояс нерухомо прикріплений до черепа. У наземних хребетних він іде від ребер та від поперечних відростків шийних хребців до внутрішньої поверхні лопатки. Крім цих парієтальних м'язів, у зв'язок з переднім поясом вступає ще один, вісцеральний м'яз - трапецієподібний (m. trapecius), який також треба віднести до вторинної мускулатури пояса. У птахів сильно розвиваються м 'язи передніх кінцівок, особливо грудний, який забезпечує роботу крил. У ссавців (мал.38) сильно розростається вздовж усієї спини широкий спинний м'яз (m. latissimus dorsi) і сильно розвивається й диференціюється грудний м'яз (m. pectoralis major et minor). Крім цього, сильно розростається й диференціюється трапецієподібний м'яз. Коракоїдно-плечовий м'яз диференціюється далі й з редукцією коракоїда частково переносить місце свого відходу на лопатку. Разом з коракоїдно-променевим м'язом він бере участь в утворенні характерного для ссавців двоголового м'яза (m.biceps brachii). Набувають великого значення зубчасті м'язи. У тварини, що піднімає своє тіло над землею, воно неначе висить між лопатками саме на зубчастих м'язах.
Мал.38. Поверхнева мускулатура тулуба та кінцівок собаки (за Мартіном). І - три-чотириголовий; 2 - двоголовий; 3 - дельтовидный; 4 - розгинач пальців; 5 -згинач пальців; б - широкий м 'яз спини; 7 -зовнішній косий м 'яз черева; 8 - грудний; 9 - прямий м 'яз черева; 10 - грудинно-ключично-сосковидний; 11 - трапецієвидний. У задній кінцівці немає вторинної мускулатури пояса, бо останній прикріплений до хребта й більш чи менш зафіксований у своєму положенні. Первинна мускулатура складається з таких найголовніших м'язів: привідні (т. adductores),гребенястий (т. pectineus), що лежать спереду, лобково-стегновий (т. pubofemoralis) - з внутрішнього боку кінцівки,грушоподібний (т. pyriformis) - ззаду. Сідничний (m. glutaeus), диференційований у ссавців на кілька м'язів, - ззаду та ззовні. М'язи, що обмежуються границями вільної кінцівки, поділяються на розгиначі (m. extensores), які виникли внаслідок диференціювання первинної спинної мускулатури, і згиначі (m. flexores) - результат диференціювання первинної черевної мускулатури. Чотириголовий м'яз (m. quadriceps femoralis) складний і йде від клубової, лобкової та стегнової кісток через коліно до великої гомілкової кістки. В її сухожилку розвивається колінна чашечка (patella). Складне диференціювання мускулатури кінцівок явно зв'язане зі зміною їх функцій при переході від водного життя до наземного. Для виконання функції рулів або весел досить було порівняно простої мускулатури, що служила для зміни положення лопаті плавця, яка є простою пружною пластинкою. З перетворенням кінцівки в складний важіль, що підтримує й пересуває тіло тварини на суші, природно, пов'язане відокремлення великої кількості м'язів, координованою дією яких обумовлюються різнономанітні рухи наземних хребетних. Мускулатура тулуба власне відступає на задній план, а м'язи кінцівок займають переважаюче положення, значною мірою покривають тулубову мускулатуру й навіть частково її витісняють. Надзвичайно важливим набуттям ссавців, зв'язаним з розвитком легеневого дихання, є діафрагма. Вона являє собою мускульну перегородку, що відділяє грудну порожнину з легенями й серцем від черевної порожнини. Перший зачаток її появляється у вигляді поперечної сполучнотканинної перегородки, яка розвивається в передній частині тіла безпосередньо позаду серця у зв'язці з задньою стінкою перикардія. Одночасно з переміщенням серця перегородка переміщається назад. У неї вростає черевна тулубова мускулатура у вигляді пари м'язових складок, що ростуть від черевної стінки тіла. У дорослої тварини або вся діафрагма є мускульною, або середина її складається зі сполучної тканини. Іннервується діафрагма нервом, який відходить від шийного сплетіння спинномозкових нервів. Така іннервація також вказує на переміщення діафрагми назад, що пов'язано з процесом відокремлення шиї в наземних тварин. Очні м'язи розвиваються з трьох (п'яти) перед вушних міотомів голови, які розпадаються на мезенхіму, що утворює потім зачатки окремих м'язів. У всіх хребетних розвиваються чотири прямі м'язи, що відходять від дна орбіти з задньої її частини й прикріпляються до верхнього, переднього, нижнього й заднього краю очного яблука, і два косі м'язи, що відходять від передньої стінки орбіти й прикріпляються до верхньої й нижньої стінок очного яблука (мал.39).
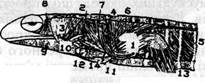
Мал.39. Очні м'язи та нерви, що їх іннервують. М 'язи: І - верхній косий; 2 - нижній косий; 3 - нижній прямий; 4 - внутрішній прямий; 5 - верхній прямий; 6 - зовнішній прямий; нерви: ІІІ - окоруховий; IV-блоковий; V-відвідний. 3 першого передщелепного міотома розвиваються три прямі м'язи - верхній (m. rectus superior), внутрішній (m. rectus internus), нижній (m. rectus inferior) і нижній косий м'яз (m. obliquus inferior). Усі вони іннервуються окоруховим нервом (третьої пари), що є нервом цього сегмента. З другого щелепного міотома, що є подвійним, розвивається один тільки верхній косий м'яз (m. obliquus superior), який іннервується блоковим нервом (четвертої пари), а з третього (також подвійного) міотома - зовнішній прямий м'яз (m. rectus externus), що іннервується відвідним нервом (шостої пари). У наземних хребетних від зовнішнього прямого м'яза відокремлюється глибша порція, що йде прямо до основи очного яблука; це м'яз, що втягує очне яблуко (m. retractor), який іннервується, згідно зі своїм походженням, відвідним нервом. При втягуванні очного яблука всередину орбіти повіки почасти пасивно змикаються над оком, особливо третя повіка, або мигальна перетинка, яка звичайно позбавлена власної мускулатури. У рептилій та птахів від дистальної частини м'яза, Що втягує очне яблуко, розвивається спеціальний м'яз мигальної перетинки. У ссавців прямі м'язи ока розміщаються тісніше навколо зорового нерва, але загалом тут є ті ж м'язи, Що й у нижчих форм. Верхній косий м'яз набирає своєрідного положення: він видовжується, місце його відходу переноситься від переднього краю орбіти вглиб останньої, однак положення його кінцевої частини фіксується біля переднього краю орбіти за допомогою сухожильного вушка, що утворює блок, через який перекидається м'яз на своєму шляху, і, незважаючи на значне переміщення місця прикріплення, діяння його залишається по суті тим же. М'яз, що втягує очне яблуко, є в більшості ссавців, але в мавп він редукується. У вищих мавп та в людини лише рідко спостерігаються його залишки у ссавців іноді від верхнього прямого м'яза відокремлюється частина волокон, яка утворює м'яз, що піднімає верхню повіку. Він іннервується окоруховим нервом. Під'язикова мускулатура розвивається за рахунок черевних відростків потиличних, а іноді й передніх тулубових міотомів, які ростуть вниз, обгинаючи ззаду зябровий апарат, і потім вперед, розміщаючись у вигляді суцільної маси під вісцеральним апаратом. У дорослої тварини тут є диференційована на окремі пучки маса поздовжніх мускульних волокон, нерідко поділена сухожилковими перегородками на окремі сегменти. Ця мускулатура являє собою безпосереднє продовження вперед черевної мускулатури тулуба, і сегментація її в нижчих хребетних є залишком колишнього поділу на міомери, однак відповідність цієї сегментації початковій звичайно втрачається. У нижчих риб під'язикова мускулатура йде від плечового пояса вперед і розщеплюється на пучки, що йдуть до окремих вісцеральних дуг. У земноводних під'язикова мускулатура становить безпосереднє продовження прямого м'яза черева. Вона розподіляється, як і в усіх інших наземних хребетних, на задню ділянку, що йде від плечового пояса та груднини до під'язикового апарату - m. sternohyoideus (від груднини) і т. omohyoideus (від пояса), і передню ділянку, що йде від під'язикового апарату до підборіддя - підборідно-під'язиковий м'яз (m. geniohyoideus). Від останнього в наземних хребетних відокремлюється і власна мускулатура язика - підборідно-язиковий м'яз (m. genioglossus), що служить для висування язика, і під'язиково-язиковий м'яз (т. hyoglossus), що служить для його втягування. Уся під'язикова мускулатура іннервується відповідно до свого походження комплексом потиличних, іноді передніх спинномозкових нервів, що утворюють так званий під'язиковий нерв (n. hypoglossus), який тільки у вищих хребетних стає типовим черепномозковим нервом (12-ої пари). Повний ряд завушних міотомів голови зберігається в дорослому стані тільки в круглоротих, де вони поділяються На спинний і черевний відділи, утворюючи надзяброву та підзяброву мускулатуру. У риб і в наземних хребетних передні завушні міотоми цілком зникають за задні, потиличні, міотоми рідко зберігаються у вигляді мало змінених залишків у ділянці потилиці, здебільшого ж утворюють разом з передніми тулубовими міотомами окрему надзяброву й підзяброву, або під'язикову мускулатуру. Надзяброва мускулатура спостерігається у вигляді системи невеликих м'язів між верхніми кінцями зябрових дуг тільки в нижчих риб. Підзяброва, або під'язикова мускулатура є, навпаки, у всіх хребетних. Вісцеральна м'язова система До вісцеральної мускулатури належать м'язи шкіри й шкірних залоз, стінок кровоносних судин, вивідних протоків сечостатевої системи, глотки, кишечника й серця. Вісцеральна мускулатура в основному гладенька, в серці й глотці - поперечносмугаста. Вона розвивається за рахунок бічних пластинок соміта й іннервується вісцеральними нервами, волокна яких виходять звичайно в складі спинних корінців і мішаних головних нервів. Поперечносмугаста вісцеральна мускулатура не може бути виділена як окрема система, бо вона є лише результатом вищого диференціювання певних ділянок гладенької мускулатури (приклад - серцевий м'яз). Частина гладенької мускулатури розвивається іноді ще за рахунок основного шару ектодерми. Так розвиваються гладенькі мускульні волокна шкіри амфібій, м'язи потових і молочних залоз ссавців, кільцевий замикач (m. sphinter pupillae) і радіальний розширювач (т. dilatator pupillae) зіниці. У птахів ця мускулатура - поперечносмугаста. Усі ці м'язи іннервуються вісцеральними нервами симпатичної системи. Гладенька мускулатура шкіри розвивається за рахунок елементів шкірного листка соміту; мускулатура кишечника - за рахунок вісцерального листка несегментованої мезодерми бічних пластинок; мускулатура сечостатевих органів і кровоносних судин - за рахунок мезенхіми, що походить з бічних пластинок, або почасти (судини шкіри) з мезенхіми самої шкіри. Мускулатура шкіри складається з волокон різного напряму, які лише рідко дають початок більш чи менш відокремленим пучкам (до піхов пір'я у птахів та до сумок волосся у ссавців). Мускулатура кровоносних судин утворена неперервним шаром кільцевих волокон, які на капілярах лягають поздовжньо. На різних внутрішніх органах спостерігаються шари кільцевих і поздовжніх волокон. Стінки кишечника оточені також внутрішнім шаром кільцевих волокон та зовнішнім шаром поздовжніх, спільним діянням яких обумовлюються перистальтичні рухи кишки. У ділянці шлунка є ще середній шар косих волокон. Уся ця мускулатура іннервується вісцеральними нервами переважно симпатичної нервової системи. Ми бачили, що гладенька мускулатура іноді стає поперечносмугастою. Це спостерігається, наприклад, у райдужній оболонці ока птаха. Те саме сталося на певній ділянці черевної кровоносної судини, в якій мускульна стінка розвинулася досить сильно і стала поперечносмугастою. Таким чином розвинулося серце хребетних. Така заміна гладенької мускулатури поперечносмугастою становить у хребетних досить рідкісне явище, але є, проте, ще ціла ділянка, де подібний процес спостерігається не як виняток, а як правило. Це - передня частина кишечника, вісцеральна мускулатура якої є поперечносмугастою у всіх хребетних і простягається назад на значну частину стравоходу. Іноді ця мускулатура розростається і на ділянку шлунка чи навіть на саму кишку, де вона утворює зовнішній мускульний покрив (лин, в'юн). Іноді навіть стінка плавального міхура набуває поперечносмугастої мускулатури (кісткові ганоїди). Особливо розвивається поперечносмугаста мускулатура в ділянці глотки, де вона вступає у зв'язок з розміщеним тут вісцеральним скелетом і стає довільною. Ця мускулатура зазнає в хребетних значного диференціювання, утворює більшість м'язів голови й становить власне вісцеральну мускулатуру, точніше вісцеральну мускулатуру вісцерального апарату. У нижчих хребетних більша її частина складається з цілком неперервної маси кільцевих волокон, що охоплюють увесь вісцеральний аппарат знизу та з боків й утворюють загальний стискач (m. constrictor superficialis). Biд цього м'яза уже в круглоротих та селахій відокремлюються окремі глибші мускульні пучки, що тісніше сполучаються з окремими вісцеральними дугами. У передній частині вісцерального апарату частина цих м'язів зазнає складнішої диференціації, особливо в круглоротих, у зв'язку з розвитком складного їх губного апарату сисного типу. У нижчих риб ще досить добре розвинений загальний стискач вісцерального апарату у вигляді шару мускульних волокон, що охоплюють увесь апарат збоку і знизу. У цьому неперервному м'язі розрізняють лише певні ділянки, наприклад, спинну частину в ділянці вісцеральних щілин і черевну, що лежить знизу (мал.40).
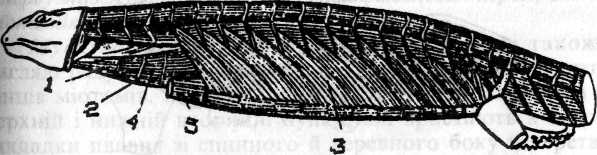
Мал. 40. Вісцеральна мускулатура акули (за Феттером). 1-м'яз, що приводить щелепу; 2-перша зяброва щілина; 3-загалышй стискач; 4-між-щелеппий м'яз; Б- м'яз, що піднімає піднебінно-квадратний хрящ; 6- бриз-кальце; 7-трапецісвидпий м яз. Далеко істотніший поділ його на ділянки іннервації, які збігаються з загальним сегментарним поділом усього апарату. Спереду, в ділянці щелепної дуги аж до бризкальця, м'яз іннервується трійчастим нервом; далі ззаду, в ділянці під'язикової дуги від бризкальця і до першої зябрової щілини, м'яз іннервується лицевим нервом; потім в ділянці першої зябрової дуги аж до другої зябрової щілини - язиково-глотковим нервом, і, нарешті, вся задня частина м'яза являє собою ділянку іннервації блукаючого нерва. У передній частині стискача спостерігається відокремлення крупних мускульних мас, що прикріпляються До обох відділів щелепної дуги. Частина, що прикріпляється до верхнього відділу щелепної дуги, лежить глибше і являє собою м'яз, який піднімає піднебінно-квадратний хрящ (in. levator palatoquadrati).Частина, що прикріпляється до нижнього відділу, являє собою м'яз, який приводить у рух нижню щелепу (m. adductor mandibulae). Ці м'язи іннервуються трійчастим нервом. Черевна частина стискача, яка лежить знизу між гілками нижньої щелепи, являє собою міжщелепний м'яз (in. intermandibularis). Передня частина останнього іннервується трійчастим нервом, а задня - лицевим. Нарешті, ззаду зябрового апарату із спинного боку спостерігається відокремлена частина загального стискача, яка невеликими пучками прикріпляється до останніх зябрових дуг, а головною своєю масою - до переднього краю спинного відділу плечового пояса. Це - трапецієподібний м'яз (m. trapezius) (мал.40). У вищих риб мускулатура вісцерального апарату трохи складніше диференційована. Відокремлюються особливі м'язи підвіска та зябрової кришки. У земноводних та в інших наземних хребетних у зв'язку з розвитком аутостилії редукувався і м'яз, що піднімає піднебінноквадратний хрящ. М'яз, що приводить нижню щелепу, розпадається на більш поверхневий жувальний м'яз (m. masseter) і глибший - висковий (т. temporalis). У вищих наземних хребетних крім цих м'язів відокремлюється ще глибша мускульна маса, що відходить від крилоподібної кістки й прикріпляється до внутрішньої поверхні нижньої щелепи. Це - крилоподібний м'яз (m. pterygoideus). Між гілками нижньої щелепи зберігається передня частина загального стискача у вигляді міжщелепного м'яза. У ділянці мускулатури лицевого нерва в наземних хребетних замість м'язів підвіска та зябрової кришки риб спостерігається досить великий м'яз, що прикріпляється до заднього кута нижньої щелепи. Це м'яз, що опускає нижню щелепу (m. depressor mandibulae). Задня частина поверхневого стискача розростається в рептилій у досить великий м'яз, що охоплює шию знизу і з боків. Це стискач шиї (m.sphinter colli). У ссавців він поділяється на глибший шар, що зберігає ту ж назву, і поверхневий шар, що розміщається безпосередньо під шкірою (m. platysma myoides). Волокна цих шарів мають косий напрям і перехрещуються між собою. Ці м'язи розростаються на всю ділянку голови й дають початок складній системі підшкірної лицевої мускулатури. У мавп і в людини ці м'язи особливо складно диференційовані й утворюють так звану мімічну мускулатуру. М'язи власне зябрового апарату з втратою зябрового дихання в наземних хребетних редукуються, але почасти зберігаються у вигляді м'язів під'язикового апарату, глотки, гортані. Трапецієподібний м'яз остаточно втрачає зв'язок із зябровими дугами і стає виключно м'язом плечового пояса. Він розростається в наземних хребетних, особливо у ссавців, де відходить не тільки від шийної та грудної ділянок хребта, а й від самого черепа аж до соскоподібного його відростка і прикріпляється не тільки вздовж гребеня лопатки та її акроміального відростка, але й до ключиці і навіть до груднини. Ця нижня частина м'яза іноді відокремлюється і дістає назву груднинно-ключично-соскоподібного м'яза (т. sternocleidomastoideus), тим часом, як верхня частина зберігає попередню назву. Іннервуюча цей м'яз задня частина блукаючого нерва відокремлюється у вищих наземних хребетних і стає самостійним нервом голови -додатковим. Отже, для хребетних тварин характерний різкий поділ усієї м'язової системи на соматичну й вісцеральну мускулатуру, різні щодо походження й іннервації. Соматична мускулатура служить спочатку тільки для пересування тварини у воді за допомогою ундуляційних згинів тіла й поділена у зв'язку з цим на ряд окремих сегментів. З утратою передньою частиною тіла своєї гнучкості та утворенням голови міомери тут, природно, редукувались за винятком тих, які набули іншої функції - очні м'язи та поздовжня мускулатура вісцерального апарату. Зі зміною способу пересування в наземних хребетних і тулубова мускулатура втрачає свою сегментацію й диференціюється на складну систему м'язів різного значення, зв'язаних почасти з осьовим скелетом та його похідними і з поясом кінцівок, які іноді виконують роль черевного преса та дихальних м'язів. Мускулатура кінцівок, що розвинулась спочатку як сегментальний утвір, втрачає поступово свою сегментацію вже в риб, але зазнає особливо складного диференціювання лише в наземних хребетних у зв'язку зі зміною функцій кінцівок. Нарешті, у вищих наземних хребетних за рахунок поверхневих шарів різних м'язів іноді розвивається ще Досить значна шкірна поперечносмугаста мускулатура.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 667; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.216.208.243 (0.016 с.) |