Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Частотное распределение фенотиповСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Каждая особь в данной популяции любого вида характеризуется присущим
только ей комплексом признаков, и можно построить графики частотного распределения этих признаков. Например, в популяции мышей величина помета может варьировать от 2 до 16 детенышей. Если мы отметим процентное соотношение каждого возможного размера помета на графике (рис. 5.2), то получим распределение частот. Если бы потомство было одинаковым и если бы у особей любого фенотипа число выживших потомков было одинаковым, тогда форма кривой частотного распределения оставалась бы неизменной из поколения в поколение. В том случае, когда изменение частотного распределения сохраняется в течение более одного поколения, можно сказать, что эволюция имеет место. Изменения могут происходить в случае, если особи, характеризующиеся одним фенотипом оставляют меньше выживших потомков, чем особи, характеризующиеся другим фенотипом. У мышей с небольшой величиной помета число выживших потомков может быть таким же, как и у мышей с большими выводками. У первых может быть большее число пометов, чем у вторых, или доля выживших может быть ниже у особей с большими пометами, чем у особей с маленькими пометами. Однако изменения в условиях обитания могут благоприятствовать появлению пометов какого-то конкретного размера таким образом, что из таких пометов выживает и размножается больше мышей. В этом случае происходит эволюция размеров выводка. Естественный отбор может по-разному действовать на распределение частот фенотипов в популяции. В результате мутаций, рекомбинаций и дрейфа генов разнообразие признаков возрастает от одного поколения к другому. Эта тенденция обычно противостоит стабилизирующему отбору, который работает сравнительно более жестко на границах распределения частот (рис. 5.3). Если давление отбора по-разному действует вдоль некоего фенотипического градиента, то имеет место направленный отбор. Это может привести к сдвигу среднего значения распределения частот или к асимметричному распределению без изменения среднего значения. Иногда возможно сильное отрицательное давление отбора по отношению к признакам, типичным для данной популяции; в этом случае отбор благоприятствует животным с отклоняющимися признаками (рис. 5.3). Этот так называемый дизруптивный отбор редок и приводит к бимодальному распределению частот. В некоторых случаях он может привести к разделению одного вида на два разных. Относительное число различных генов может изменяться случайным образом от поколения к поколению и без действия естественного отбора. Этот эффект, известный как случайный дрейф генов (Wright, 1921), сильнее всего проявляется в небольшой популяции. На рис. 5.4 показано, как чисто случайно может элиминироваться в маленькой популяции конкретный генотип. Изменчивость, вызванная случайными отклонениями, иногда при-
Рис. 5.4. Схематическое изображение дрейфа генов. В каждом поколении особь с обусловливающим серую или белую окраску генотипом производит две такие же особи. Половина молодых птиц погибает, и величина популяции остается постоянной. Однако гибель молодых птиц определяется чистой случайностью и не зависит от генотипа. Частота каждою генотипа флуктуирует случайным образом, пока серые особи не вымирают и белые не закрепляются в популяции. (По Futuyma, 1979.)
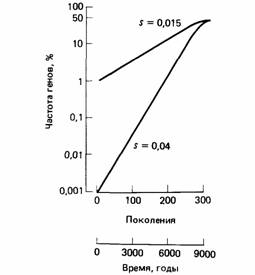
водит к необычному соотношению генотипов, особенно в небольших изолированных популяциях (рис. 5.5). Например, в некоторых изолированных альпийских деревнях частота случаев альбинизма в 10 раз выше, чем обычно. Известно, что в других изолированных селениях значительно чаще встречаются цветовая слепота, некоторые типы умственной отсталости или глухонемота. Скорость изменения частоты генов, на которую влияет естественный отбор, зависит от относительной приспособленности различных генотипов. Относительная приспособленность генотипа отражает различия в приспособленности между данным генотипом и другими, представленными в той же популяции. Если приспособленность генотипа с наиболее высокой скоростью увеличения частоты принять за 1,00, то относительная приспособленность другого генотипа со скоростью увеличения частоты, скажем, 85% была бы равна 0,85. Разница 0,15 называется коэффициентом отбора, направленного против худшего генотипа. Коэффициенты отбора можно использовать для подсчета скорости изменения фенотипа в популяции. Интересный пример можно взять из истории одомашнивания животных около 10 000 лет назад. До появления домашних животных единственным источником молочной пищи для младенцев было грудное молоко, которое они получали в течение первых лет жизни. В желудке ребенка вырабатывается фермент лактаза, который необходим для расщепления молочного сахара - лактозы. На более поздних стадиях онтогенеза у многих людей синтез этого фермента прекращается, и во многих современных человеческих популяциях у взрослых этот фермент отсутствует совершенно. Если человек, у которого не вырабатывается лактаза, пьет молоко, то у него начинаются тошнота, рвота и боли в животе. Сохранение синтеза лактазы во взрослом состоянии - наследственная черта, которая, как полагают, появилась в результате мутации, выгодной в популяциях, для которых молоко постоянно входит в состав диеты. По-видимому, эта мутация была редкой до появления домашних животных, когда люди всецело жили охотой и собиранием растений. Таким образом, в некоторых популяциях частота генотипов, контролирующих переносимость лактозы, увеличилась, по-видимому, от первоначально очень низкой до современной высокой, как у жителей северной Европы и других регионов, где потребляют молоко в больших количествах. Предположим, что время смены поколений у человека в среднем составляет 30 лет. Следовательно, с начала одомашнивания животных сменилось примерно 300 поколений. В течение этого периода доля взрослых, у которых вырабатывается лактаза, среди жителей северной Европы возросла почти от нуля до 75%. Допустим, что этот признак определяется одним геном, тогда его частота должна была возрасти от 0,001% до 50%, необходимых
дляполучения данной частоты генотипа в 75%. Можно рассчитать, что для полученияэтих изменений был бы необходим коэффициент отбора, равный 0,04. Если первоначальная частота гена, контролирующего синтез лактазы, была 1,0%, то достаточно было бы коэффициента 0,015 (рис. 5.6). Это означает, что относительно быстрые эволюционные изменения возможны, даже если разница в приспособленности между двумя аллелями меньше 5%. Можно подсчитать (Futuyma, 1979, с. 314-316), что скорость увеличения частоты аллелей с благоприятными признаками пропорциональна коэффициенту отбора и частоте в популяции как благоприятных, так и вредных аллелей. Поэтому скорость генетических изменений высока только тогда, когда оба гена часто встречаются в популяции. Частота появления новых мутаций сначала возрастает очень медленно. Более того, полное замещение одного аллеля другим требует очень большого промежутка времени, поскольку этот процесс замедлен в начале, когда благоприятный аллель редок, и в конце, когда становится редким неблагоприятный аллель. Вредные рецессивные аллели редко полностью элиминируются естественным отбором, так как они проявляются в фенотипе только в гомозиготном состоянии. Поэтому в природных популяциях содержится много вредных рецессивных генов, которые редко проявляются и образуют своего рода генетическое хранилище. Скорость эволюции отдельного гена зависит от его фенотипического проявления, от его частоты в популяции и коэффициента отбора. Например, частота редкого благоприятного доминантного гена возрастает гораздо более резко, чем частота редкого благоприятного рецессивного гена (рис. 5.7). Однако доминантный ген окончательно фиксируется в популяции (замещая альтернативный аллель) медленнее, чем рецессивный, так как оставшиеся немногие рецессивные аллели существуют в гетерозиготной форме и, таким образом, защищены от действия естественного отбора. Нет ни одного вида животных, который существовал бы отдельно в отсутствие других форм жизни, а процесс адаптации одного вида может изменить условия существования других видов. Это вызывает компенсаторную адаптацию у видов, подвергшихся такому влиянию. Так, если какой-либо вид птиц адаптируется к дефи- циту основного вида корма путем расширения диеты и включения в нее новых видов насекомых, то в условия обитания популяции этих насекомых вводится новый фактор отбора. Поскольку такие взаимосвязи можно проследить в каждой экосистеме, мы всегда должны оценивать поведение животного, учитывая комплекс экологических взаимоотношений. Экология и поведение При изучении поведения животных в природных условиях важно понимать влияние последствий поведения на способность животного к выживанию. Последствия конкретного вида деятельности зависят в основном от непосредственных условий обитания животных. В условиях, к которым животное хорошо адаптировано, последствия того или иного вида деятельности могут быть полезными. Та же деятельность, осуществляемая в других условиях, может быть вредной. Чтобы представить, как в процессе эволюции сформировалось поведение животных, нам надо понять адаптацию животных к окружающей среде. Экология - этоотрасль естествознания, которая изучает взаимосвязи животных и растений с окружающей их природной средой. Она имеет отношение ко всем аспектам этих взаимосвязей, в том числе к потокам энергии через экосистемы, физиологии животных и растений, структуре популяций животных и их поведению и т.д. Помимо получения точных знаний о конкретных животных эколог стремится понять общие принципы экологической организации, и здесь мы рассмотрим некоторые из них. В процессе эволюции животные адаптируются к конкретным условиям окружающей среды, или местообитанию. Местообитания обычно характеризуют, описывая их физические и химические признаки. Тип растительных сообществ зависит от физических свойств среды, таких, как почва и климат. Растительные сообщества обеспечивают разнообразие возможных местообитаний, которые используются животными. Ассоциация растений и животных в совокупности с конкретными условиями природного местообитания образует экосистему. На земном шаре насчитывают 10 основных типов экосистем, называемых биомами. На рис. 5.8 показано распределение главных наземных биомов мира. Существуют также морские и пресноводные биомы. Например, такой биом, как саванны, занимает большие области Африки, Южной Америки и Австралии и представляет собой травянистые равнины с редко растущими на них деревьями в тропических и субтропических областях земного шара. Для саванн типично наличие сезона дождей. На верхнем конце диапазона распределения осадков саванна постепенно сменяется тропическими лесами, а на нижнем конце - пустынями. В африканской саванне преобладают акации, в южно-американской - пальмы, а в австралийской - эвкалипты. Характерной чертой африканской саванны является большое разнообразие растительноядных копытных, которые обеспечивают существование разнообразных хищников. В Южной Америке и Австралии те же самые ниши заняты другими видами. Совокупность животных и растений, населяющих конкретное местообитание, называется сообществом. Виды, образующие сообщество, делятся на продуценты, консументы и редуценты. Продуценты — это зеленые растения, которые улавливают солнечную энергию и превращают ее в химическую. Консументы - этоживотные, которые поедают растения или травоядных животных и таким образом косвенно в энергетическом отношении зависят от растений. Редуценты - этообычно грибы и бактерии, разлагающие отмершие остатки животных и растений до веществ, которые снова могут использоваться растениями. Ниша - этороль животного в сообществе, определяемая его взаимосвязями как с другими организмами, так и с физическим окружением. Так, травоядные животные обычно питаются растениями, а травоядных животных в свою очередь поедают хищники. Виды, занимающие, данную нишу, различны в разных частях земного шара. Например, нишу мелких растительноядных в зонах умеренного климата в
Рис. 5.8. Распределение основных наземных биомов мира. Северном полушарии занимают кролики и зайцы, в Южной Америке - агути и вискаши, в Африке - даманы и белоногие хомячки, а в Австралии - валлаби. В 1917 г. американский эколог Гриннелл (Grinnell) впервые выдвинул теорию ниш, основываясь на исследовании калифорнийского пересмешника (Toxostoma redivivum) - птицы, которая гнездится в густой листве на высоте одного-двух метров над землей. Местоположение гнезда - это одна из характеристик, с помощью которой можно описать нишу животного. В горных областях необходимая для гнездования растительность имеется только в экологическом сообществе, называемом чапаралъ. Местообитание пересмешника, описываемое физическими характеристиками окружающей среды, определяется частично и реакцией популяции пересмешника на ситуацию, складывающуюся в нише. Так, если высота гнезда над землей - решающий фактор спасения от хищников, то в популяции будет наблюдаться сильная конкуренция за места для гнезд на оптимальной высоте. Если же этот фактор был бы не столь решающим, то большее число особей смогли бы строить гнезда в других местах. На условия обитания в данной нише воздействует также и конкуренция со стороны других видов за места для гнезд, пищу и пр. Местообитание калифорнийского пересмешника определяется частично ситуацией с нишами, распределением других кустарниковых видов, характерных для чапараля, и плотностью популяции самого пересмешника. Ясно, что если плотность его невелика, птицы гнездятся только в наилучших местах, и это оказывает влияние на местообитание вида. Таким образом, суммарные взаимосвязи пересмешника с условиями местообитания, которое часто обозначается термином экотоп, являются результатом сложных взаимодействий характерных особенностей ниши, местообитания и популяции. Если животные разных видов используют одни и те же ресурсы, характеризуются некоторыми общими предпочтениями или пределами устойчивости, то мы говорим о перекрывании ниш (рис. 5.9). Перекрывание ниш приводит к конкуренции,
особенно когда ресурсы недостаточны. Принцип конкурентного исключения гласит, что два вида с одинаковыми нишами не могут существовать в одном месте в одно и то же время при ограниченных ресурсах. Из этого следует вывод, что если два вида сосуществуют, то между ними должны быть экологические различия. В качестве примера рассмотрим взаимоотношение ниш у группы видов птиц, «обирающих листья», которые питаются на дубах гористого побережья в центральной Калифорнии (Root, 1967). Эту группу, называемую гильдией, составляют виды, которые используют одни и те же природные ресурсы одинаковым образом. Ниши этих видов в значительной степени перекрываются, и поэтому они конкурируют друг с другом. Преимущество концепции гильдии состоит в том, что в данном случае анализируются все конкурирующие виды данного участка независимо от их таксономического положения. Если рассматривать диету этой гильдии птиц как элемент их среды обитания, то следует сказать, что большая часть этой диеты должна состоять из членистоногих, собираемых с листьев. Это произвольная классификация, поскольку любой вид может быть членом более чем одной гильдии. Например, равнинная синица (Parus inornatus) относится к гильдии птиц, обирающих листья, на основе ее пищедобывательного поведения; кроме того, она является также членом гильдии птиц, гнездящихся в дуплах в силу гнездовых требований.
Хотя в данном случае пять видов птиц питаются насекомыми, каждый вид ловит насекомых, отличающихся по размерам и таксономическому положению. Таксономические категории насекомых, поедаемых этими пятью видами, перекрываются, однако каждый вид специализируется на определенном таксоне. Точно так же на рис. 5.10 показано, что размеры жертв полностью перекрываются, но средние значения и дисперсии у них разные, по крайней мере в некоторых случаях. Рут (Root, 1967) обнаружил также, что для птиц этих видов характерны три типа пищедобывательного поведения: 1) собирание насекомых с поверхности листьев, когда птица передвигается по твердому субстрату; 2) собирание насекомых с поверхности листьев парящей птицей; 3) ловля летающих насекомых. Доля времени, которая затрачивается каждым видом на тот или иной способ добывания пищи, показана на рис. 5.11. Этот пример отчетливо демонстрирует процесс экологической специализации в поведении. Поведение каждого вида влияет на поведение других видов таким образом, что у членов этой гильдии развиваются все возможные типы пищедобывательного поведения и используются все виды жертвы. Конкуренция часто приводит к доминированию одного вида; это выражается в том, что доминантные виды имеют преимущество в использовании таких ресур- сов как пища, пространство и убежища (Miller, 1967; Morse, 1971). Исходя из теории, следует ожидать, что вид, который становится в подчиненное положение по отношению к другому виду, должен был быизменить использование ресурсов таким образом, чтобы уменьшить перекрывание с доминантным видом. Обычно при этом субординантный вид уменьшает использование некоторых ресурсов, уменьшая таким образом ширину ниши. В некоторых случаях субординантный вид может расширить нишу, включив неиспользовавшиеся ранее ресурсы либо путем подчинения других видов в смежных нишах, либо путем более полного использования фундаментальной ниши. Если субординантный вид и выживает в конкуренции с доминантным видом, значит, его основная ниша шире, чем ниша доминантного вида. Такие случаи отмечены у пчел и черных дроздов нового света (Orians, Willson, 1964). Поскольку приоритет в использовании ресурсов принадлежит доминантным видам, субординантные виды могут быть исключены из пространства ниши, когда ресурсы ограничены, количество их непредсказуемо, а поиски пищи требуют значительных усилий; и все это значительно снижает приспособленность субординантного вида в области перекрывания. В таких случаях можно ожидать, что субординантные виды будут подвергаться значительному давлению отбора и изменят свои фундаментальные ниши либо путем специализации, либо выработав устойчивость к более широкому диапазону физических условий среды обитания. Адаптивность поведения Натуралисты и этологи обнаружили многочисленные примеры удивительных способов, которыми животные оказываются в совершенстве приспособленными к условиям среды обитания. Трудность в объяснении такого рода поведения животных состоит в том, что оно кажется убедительным только потому, что различные детали и наблюдения слишком хорошо увязываются друг с другом; иными словами, хорошая история может показаться убедительной просто потому, что это хорошая история. Это не означает, что хорошая история не может быть правдивой. В любом правильном объяснении поведенческой адаптации разнообразные детали и наблюдения и в самом деле должны быть подогнаны друг к другу. Проблема заключается в том, что биологи, как ученые, должны оценивать данные, а хорошее описание - не всегда хорошие данные.Как и в суде, данные должны быть более чем обстоятельными и должны нести в себе некоторые элементы независимого подтверждения. Один из способов получить данные, свидетельствующие об адаптивности поведения, - сравнить родственные виды, которые занимают различные местообитания. Классический пример такого подхода - работа Эстер Каллен (Ester Cullen, 1957) по сравнению особенностей гнездования у гнездящейся на скалах моевки (Rissa tridactyla) и у гнездящихся на земле чаек, таких, как обыкновенная (Lams ridibundus) исеребристая (Lams argentatus). Моевка гнездится на скальных уступах, недоступных для хищников, и, по-видимому, эволюционировала от гнездящихся на земле чаек в результате давления хищников. Моевки унаследовали некоторые черты чаек, гнездящихся на земле, такие, как частично маскировочная окраска яиц. Яйца птиц, гнездящихся на земле, обычно хорошо маскируются для защиты от хищников, но у моевок окраска яиц не может выполнять такую функцию, поскольку каждое гнездо отмечено бросающимся в глаза белым пометом. Взрослые и молодые чайки, гнездящиеся на земле, аккуратны и избегают дефекации вблизи гнезда, чтобы не показывать его местонахождение. Таким образом, представляется наиболее вероятным, что маскировочная окраска яиц моевок - свидетельство того, что их предки гнездились на земле. Каллен (Cullen, 1957) изучила размножающуюся колонию моевок на Фарнских островах недалеко от восточного побережья Соединенного Королевства, где они гнездятся на очень узких уступах скал. Она установила, что на их яйца не нападают ни наземные животные, такие, как крысы, ни птицы, такие, как серебрис-
тые чайки, которые часто хищничают на яйцах птиц, гнездящихся на земле. Моевки питаются в основном рыбой и не пожирают яйца и птенцов из соседних гнезд, как это часто делают чайки, гнездящиеся на земле. Моевки, по-видимому, уже утратили большую часть адаптации, защищающих других чаек от хищников. Например, они не только не маскируют гнездо, - они также редко издают крики тревоги и не нападают всем скопом на хищников. У моевок много специальных адаптации к гнездованию на скалах. У них легкое тело и сильные пальцы и когти, которые позволяют цепляться за уступы, слишком маленькие для других чаек. В сравнении с чайками, гнездящимися на земле, у взрослых моевок есть ряд поведенческих адаптации к скальному местообитанию. Их поведение при драках ограничено строгими рамками стереотипа в сравнении с родственниками, гнездящимися на земле (рис. 5.12). Они строят довольно искусные чашеобразные гнезда, используя для этого ветки и ил, тогда как гнездящиеся на земле чайки строят рудиментарные гнезда из травы или морских водорослей, не используя ил как цемент. Птенцы моевки отличаются от птенцов других чаек множеством признаков. Например, они остаются в гнезде в течение более длительного периода и проводят большую часть времени, повернувшись головой к скале. Они выхватывают отрыгиваемую пищу прямо из глотки родителей, тогда как большинство чаек подбирают ее с земли, куда она выбрасывается взрослыми. Птенцы гнездящихся на земле чаек при испуге убегают и прячутся, а молодые моевки остаются в гнезде. Для птенцов чаек характерны криптические окраска и поведение, тогда как у птенцов моевок этого нет. Сравнение видов может пролить свет на функциональное значение того или иного типа поведения следующими способами: когда какой-либо тип поведения наблюдается у одного вида, но не наблюдается у другого, это может быть связано с различиями в способах действия естественного отбора на эти два вида. Например, серебристые чайки удаляют яичную скорлупу вблизи гнезда, для того чтобы поддерживать маскировку гнезда, поскольку внутренняя белая поверхность яичной скорлупы хорошо заметна. Данные, поддерживающие эту гипотезу, получены при наблюдениях за моевками, которые не убирают скорлупу. Как мы уже видели, на гнезда моевок хищники не нападают и их гнезда и яйца не маскируются. Если удаление яичной скорлупы служит в основном для поддержания маскировки гнезда, тогда вряд ли мы обнаружим это у моевок. Однако, если это служит другим целям, например предупреждению болезней, тогда можно ожидать, что такое поведение будет наблюдаться у моевок. Моевки обычно содержат гнездо очень чистым и выбрасывают из него любые посторонние предметы. Серебристые чайки обычно этого не делают. Приведенные выше данные получат дополнительное подкрепление, если мы сможем показать, что у других родственных видов, находящихся под таким же давлением отбора, появляются сходные адаптации. Один такой пример приводит Хейлман (Hailman, 1965), изучавший гнездящуюся на скалах вилохвостую чайку (Lams furcatus) на Галапагосских островах. Хейлман изучал различные виды поведения, которые определяются возможностями предотвратить опасность паления со скал. Вилохвостые чайки гнездятся и не на таких крутых скалах, как моевки, и не так высоко над землей. Таким образом, можно было бы ожидать, что соответствующие адаптации вилохвостых чаек будут промежуточными между адаптациями моевок и типичных гнездящихся на земле чаек. Вилохвостые чайки подвергаются более частым нападениям хищников, чем моевки, и Хейлман обнаружил некоторые черты поведения, которые, очевидно, определяются этим различием. Например, как упоминалось выше, птенцы моевок испражняются на край гнезда, таким образом делая его весьма заметным. Птенцы же вилохвостой чайки испражняются за кромкой этого края. Он установил, что по ряду признаков, также связанных с интенсивностью хищничества, вилохвостые чайки занимают промежуточное положение между моевками и другими чайками. Таким образом Хейлман оценил те поведенческие признаки вилохвостых чаек, которые являются адаптациями к наличию имеющегося пространства для гнездования и наличию мест для гнезд и гнездового материала. Затем он решил оценить данные, на которых была основана гипотеза Каллен (Cullen, 1957) о том, что характерные черты моевок - это результат давления отбора, сопровождающего гнездование на скалах. Он выбрал 30 признаков вилохвостой чайки и разделил их на три группы в зависимости от степени сходства с поведением моевок. Взятое в целом, это сравнение подтверждает гипотезу Каллен, что особые черты моевок - это результат действия отбора, который сопровождает гнездование на скалах. Работа Крука (Crook, 1964) на почти 90 видах ткачиков (Ploceinae) - другой пример такого сравнительного подхода. Эти маленькие птицы распространены по всей Азии и Африке. Несмотря на внешнее сходство, различные виды ткачиков заметно различаются по социальной организации. Некоторые из них защищают большую территорию, на которой строят замаскированные гнезда, тогда как другие гнездятся колониями, в которых гнезда хорошо заметны. Крук обнаружил, что виды, живущие в лесах, ведут одиночный образ жизни, питаются насекомыми, гнезда маскируют на большой охраняемой территории. Они моногамны, половой диморфизм выражен слабо. Виды, обитающие в саванне, обычно семеноядны, живут группами, гнездятся колониально. Они полигамны, причем самцы окрашены ярко, а самки - тускло. Крук считал, что, поскольку пищу в лесу трудно найти, необходимо, чтобы оба родителя выкармливали птенцов, а для этого родители должны держаться вместе на протяжении сезона размножения. Плотность насекомых, которыми питаются лесные птицы, невелика, поэтому пара птиц должна защищать большую территорию, чтобы обеспечить соответствующее снабжение птенцов пищей. Гнезда хорошо замаскированы, а взрослые птицы тускло окрашены, чтобы во время их посещения гнезда хищники не могли раскрыть его местонахождение. В саванне семян может быть много в одних местах и мало в других, что является примером пятнистого распределения пищи. Отыскивание пищи в таких условиях более эффективно, если птицы образуют группы, чтобы вести поиски в широкой области. Места для гнездования, защищенные от хищников, в саванне редки, поэтому множество птиц гнездится на одном дереве. Гнезда объемистые, чтобы обеспечить защиту от солнечного тепла, поэтому колонии хорошо заметны. Для защиты от хищников гнезда обычно строят высоко на колючих акациях или других подобных деревьях (рис. 5.13). Самка сама в состоянии прокормить потомство,
поскольку пищи сравнительно много. Самец почти не участвует в этом и ухаживает за другими самками. Самцы конкурируют за места для гнезд внутри колонии, и преуспевшие в этом могут каждый привлечь нескольких самок, в то время как другие самцы остаются холостыми. В колониальном поселении ткачиков (Textor cucullatus), например, самцы воруют друг у друга гнездовой материал. Поэтому они вынуждены постоянно находиться около гнезда, чтобы охранять его. Для привлечения самок самец устраивает сложное «представление», свешиваясь из гнезда. Если самец добивается успеха в ухаживании, самка входит в гнездо. Такое привлечение внимания к гнезду типично для колониальных ткачиков. Совсем по-другому проходит ритуал ухаживания у видов птиц, живущих в лесу, у которых самец выбирает самку, ухаживает за ней на заметном расстоянии от гнезда и затем ведет к гнезду. Сравнительный подход оказался плодотворным методом при изучении взаимосвязи между поведением и экологией. С помощью этого метода изучались птицы (Lack, 1968), копытные (Jarman, 1974) и приматы (Crook, Gartlan, 1966; Glutton-Brock, Harvey, 1977). Некоторые авторы (Clutton-Brock, Harvey, 1977; Krebs, Davies, 1981) высказывают критические замечания в адрес сравнительного подхода, тем не менее он предоставляет удовлетворительные данные, касающиеся эволюционных аспектов поведения при условии, что приняты соответствующие меры, чтобы избежать подмены понятий и замыкающихся друг на друга доказательств. Хейлман (Hailman, 1965) считает сравнительный метод подходящим только в тех случаях, когда сравнение двух популяций животных позволяет сделать выводы относительно третьей популяции, которая еще не изучена к тому времени, когда эти выводы формулируются. В этом случае гипотезу, сформулированную в результате сравнительного исследования, можно проверить независимо без использования данных, полученных в результате этого исследования. Нетрудно понять, что если взаимосвязанные различия в поведении и экологии существуют между двумя популяциями, то этого еще недостаточно, чтобы говорить о том, что в данных признаках отражается давление отбора, возникающее в результате различий в условиях обитания этих двух популяций. Различий, возникающих вследствие смешения переменных или вследствие сравнения несоответствующих таксономических уровней, можно избежать тщательным статистическим анализом (Clutton-Brock, Harvey, 1979; Krebs, Davies, 1981). ДЛЯ ЗАПОМИНАНИЯ 1. Доказательства эволюции путем естественного отбора можно получить, сравнивая современные виды друг с другом и с их ископаемыми предками, а также изучая последствия географической изоляции. Мы располагаем несколькими примерами естественного отбора, идущего в настоящее время. 2. Частоты распределения фенотипов могут меняться в результате давления отбора, и скорость этого изменения зависит частично от генетической структуры популяции, а частично от проявления данного признака в фенотипе. 3. Основные типы взаимодействия между животными различных видов - хищничество и конкуренция. Для того чтобы добывать пищу, каждый вид занимает особую нишу. Виды со сходными нишами в одних местообитаниях конкурируют в борьбе за пищу и могут конкурировать также в борьбе за другие ресурсы, такие, как места для гнезд. Два вида не могут занимать одну и ту же нишу в одном и том же местообитании. 4. Сравнение близкородственных видов, живущих в различных местообитаниях, часто может выявить такие аспекты поведения, которые особенно важны в приспособлении животного к условиям окружающей среды. Рекомендуемая литература Futuyma D.J. (1979) Evolutionary Biology, Sinauer Associates, Sunderland, Massachusetts. Dawkins R. (1976). The Selfish Gene, Oxford University Press, Oxford.
|
||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 624; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.015 с.) |