Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Хеморецепция и терморецепцияСодержание книги
Поиск на нашем сайте Хеморецепцией называется способность распознавать химические вещества и определять их концентрацию. Ею обладают даже очень примитивные формы живого. Собственно говоря, любую нервную клетку можно считать хеморецептором, так как она специфически реагирует на вещества, выделяемые другими нервными клетками. Механизмы хеморецепции включают распознавание специфических молекул рецепторными участками на клеточных мембранах. Не совсем ясно, основано ли это на химическом действии, на форме молекулы или на том и другом одновременно. Например, мы не знаем, какая общая черта сахара и сахарина делает оба этих вещества сладкими для падальных мух, крыс, обезьян и человека. Хеморецепторы делятся на экстероцепторы и интероцепторы. Первые реагируют на присутствие химических веществ во внешней среде, а вторые - на соединения, циркулирующие в жидкостях тела, в частности на окись углерода, питательные вещества и гормоны. И вкус, и обоняние зависят от хеморецепторов. В традиционном понимании обоняние различает низкие концентрации веществ, содержащихся в воздухе, а вкус возникает от прямого соприкосновения с относительно высокими концентрациями химических веществ. Однако в обоих случаях на рецептор действуют растворенные вещества; у некоторых животных, например у водных обитателей, разделить эти чувства трудно. Тем не менее у многих животных имеется неврологическое различие, которое состоит в том, что одни нервы связаны с обонянием, т. е. обнаружением низких концентраций, а другие проводят вкусовые сигналы от разных рецепторов, специфически реагирующих на относительно концентрированные химические вещества. Так, у падальной мухи хеморецепторы антенн распознают малые количества веществ в воздухе, а хеморецепторы на лапках способны обнаруживать соль, сахар и чистую воду. У позвоночных вкусовые сигналы передаются по лицевому (VII) и языкоглоточному (IX) черепномозговым нервам, а обонятельные - по обонятельному нерву (1). Одна из наиболее исследованных обонятельных систем у беспозвоночных связана с восприятием феромона ухаживания у тутового шелкопряда (Bombyx mori). Феромон - это соединение или смесь соединений, выделяемая организмом в окружающую среду и вызывающая специфическую поведенческую или физиологическую реакцию у воспринимающего организма того же вида. Таким образом, феромоны - это химические посредники, участвующие, вероятно, в самой примитивной форме общения животных. Первым химически идентифицированным половым феромоном был феромон тутового шелкопряда; это вещество (многоатомный спирт) получило название «бомбикола». Его секретируег абдоминальная железа самок, которые таким способом могут привлекать самцов на расстоянии нескольких километров. Химическое строение бомбикола известно, он был синтезирован, испытывался на самцах этого вида или применялся для стимуляции изолированных антенн, к рецепторам которых были подведены электроды (Schneider, 1969; Payne, 1974). Полную физиологическую реакцию вызывает только бомбикол, а более слабый ответ возникает и на некоторые очень близкие к нему вещества. Это говорит о большой специфичности. Еще примечательнее то, что полную реакцию может вызвать всего одна молекула бомбикола (Kaissling, Priesner, 1970). Поведенческий ответ самца шелкопряда состоит в том, что он летит против ветра, ориентируясь по молекулам бомбикола и, найдя самку, спаривается с ней. Такой тип ориентации на химический стимул очень распространен у насекомых. Насекомые обладают разными типами обонятельных сенсилл (рис. 12.1). На их поверхности обычно находятся многочисленные мелкие поры, которые заканчиваются трубочками, наполненными жидкостью. Дендриты рецепторных клеток заходят в сенсиллы, а их аксоны идут прямо в мозг. Взвешенные в воздухе молекулы феромона попадают в поры сенсиллы и через них проникают в заполненную жидкостью внутреннюю часть, где приходят в соприкосновение с рецепторной мембраной. У позвоночных обонятельные рецепторы представляют собой сенсорные нейроны первого порядка, дендриты которых в виде ресничек погружены в слизистый слой (рис. 12.2). Аксоны этих нейронов идут к обонятельной луковице, где образуют синапсы с нейронами второго порядка, аксоны которых образуют обонятельный тракт, входящий в передний мозг. Вкусовые рецепторы состоят из чувствительных клеток, которые обычно расположены группами, называемыми сосочками. Чувствительные клетки плотно прилегают к сенсорным нервным волокнам, которые остаются неизменными, тогда как вкусовые клетки каждые несколько дней замещаются новыми. У млекопитающих четыре основных вкусовых ощущения - кислого, горького,
соленого и сладкого. У людей к каждому из них чувствительны разные части языка. Кислое воспринимается боковыми сторонами ближе к корню, горькое - самой задней областью, соленое - боковыми сторонами ближе к переднему краю, а сладкое -кончиком языка (рис. 12.3). Вкусовые качества пищи зависят одновременно от ее вкуса и запаха. Это легко показать, попросив испытуемого определить с закрытыми глазами разницу между мелкими кусочками яблока и лука, положенными ему на язык. Большинство людей легко их различают, если имеют возможность дышать через нос, но не способны сделать это с зажатым носом. Сделано много попыток выделить основные запахи по примеру четырех основных вкусовых ощущений. Наиболее распространенная классификация, приводимая в табл. 12.1, принадлежит Дж. Эймуру (J. Amoore, 1963). Еще будучи студентом
Оксфордского университета. Эймур заметил большие различия в химическом строении одинаково пахнущих веществ. Он исследовал более 600 органических соединений с детально описанными запахами и в 1952 г. опубликовал свою стереохимическую теорию обоняния, в которой все они охарактеризованы на основе семи первичных запахов, перечисленных в табл. 12.1. Теория Эймура классифицирует химические вещества по форме и величине их молекул. Он постулирует существование семи основных типов рецепторов, характеризующихся особыми рецепторными участками, в которые могут входить молекулы определенной формы. Так, молекулы веществ с камфарным запахом приблизительно сферические, а у веществ с мускусным запахом они имеют форму дисков. Теория Эймура получила некоторое подтверждение в дальнейших исследованиях (Amoore, 1964), но все же остается противоречивой. К хеморецепции несколько близка терморецепция, которая, вероятно, имеется у большинства животных, но изучена у относительно немногих. Чувствительные к температуре нервные окончания обнаружены у разных насекомых. Например, у таракана Periplaneta терморецепторы на лапках воспринимают температуру почвы, а на антеннах - температуру воздуха. У рыб терморецепторы находятся в коже, боковой линии и головном мозгу, что делает их очень чувствительными к изменениям температуры. Показано, что сомы реагируют на перепады температуры менее 0.1C. Многие пресмыкающиеся обладают хорошо развитой температурной чувстви- тельностью и терморецепторами в головном мозгу и коже. У ямкоголовых змей на голове имеются так называемые лицевые ямки, чувствительные к инфракрасному излучению и устроенные так, что животное способно к дирекциональной температурной чувствительности тела. Покровы птиц, как полагают, бедны терморецепторами, за исключением языка и клюва некоторых видов. У голубей (Columbidae) терморецепторы головного мозга влияют на поведение и расположение оперения, а другие - в спинном мозгу - управляют дрожью и одышкой. У млекопитающих специальные тепловые и холодовые рецепторы распределены по коже, причем тепловые обычно залегают глубже Холодовых. Имеются также рецепторы в глубине тела (например, в венах), которые могут вызывать дрожь, даже если температура кожных и мозговых рецепторов не изменяется. Терморецепторы спинного мозга влияют на дрожь, одышку и кровоток, причем эти же функции дублируются терморецепторами гипоталамуса. У млекопитающих в целом наблюдаются самые развитые формы терморегуляции. Их головной мозг получает сигналы от многих частей тела: их интеграция ведет к соответствующему включению различных согревающих и охлаждающих механизмов. Табл. 12.1. Первичные запахи с химическими и бытовыми примерами (по Amoore, 1963)
Механорецепторы и слух Звук возникает вследствие ничтожных изменений давления, вызываемых источником колебаний в воздухе или в воде. Звуковые рецепторы в принципе представляют собой механорецепторы с быстрым восстановлением чувствительности, что делает их восприимчивыми к колебаниям. У множества членистоногих описаны чувствительные к вибрации волоски и рецепторы в суставах конечностей. У падальных мух в некоторых суставах антенн рецепторы, называемые джонстоновыми органами, могут реагировать на колебания частотой до 500 Гц. У комаров орган такого же типа сигнализирует о направлении звука. Гладыш Notonecta локализует добычу по волнам, распространяющимся на поверхности воды. Рецепторы, расположенные на его лапках, максимально чувствительны к колебаниям частотой 100-150 Гц. Пауки отличают живую добычу от мертвой по вибрации паутины. Некоторые позвоночные обладают механорецепторами, способными воспринимать колебания субстрата. Такие рецепторы найдены в коже змей и в суставах ног у кошек и уток (Prosser, 1973). Слуховые системы животных, несмотря на разнообразие, имеют некоторые общие черты. Так, например, их периферическая часть преобразует звуковое давление в колебательное движение. Рецепторы превращают это движение в нервные импульсы, которые декодируются ЦНС. Один из самых простых типов периферических устройств известен у бабочек совок. У них имеются два «уха» (тимпанальных органа), каждое из которых состоит просто из тимпанальной мембраны по бокам груди и двух рецепторных клеток, погруженных в полоску соединительной ткани (рис. 12.4). Это поразительно простое «ухо» позволяет бабочкам слышать ультразвуковые сигналы охотящихся летучих мышей. В серии изящных опытов Редер (Roeder, 1963, 1970) показал, как это происходит. Один рецептор, называемый клеткой A1, чувствителен к слабым звукам и реагирует на сигналы летучих мышей на расстоянии около 30 м, т. е. слишком далеким для того, чтобы мышь обнаружила бабочку. Частота импульсов, идущих от клетки A1, пропорциональна силе звука, и поэтому бабочка может определить, приближается ли летучая мышь. Сравнивая время прихода и интенсивность стимула вобоих органах слуха, бабочка узнает о направлении полета мыши. Разница возникает потому, что тело бабочки заслоняет звук от одного «уха» больше, чем от другого. Может быть также определена относительная высота полета летучей мыши: когда она выше насекомого, звук, приходящий к тимпанальным органам, ритмически прерывается взмахами крыльев бабочки; когда мышь находится ниже бабочки, этого не происходит.
Клетки А1 заранее предупреждают бабочку о приближении летучей мыши и могут позволить ей улететь до того, как мышь ее обнаружит. Удаляясь по прямой, бабочка становится наименьшей мишенью, потому что ее крылья обращены к мыши краями, а не плоскостью; она добивается этого, просто разворачиваясь так, чтобы уравнять звук, достигающий обоих «ушей». Но если летучая мышь обнаруживает бабочку, та не может спастись от нее, просто улетев, так как летучая мышь движется гораздо быстрее. Поэтому бабочка уклоняется от хищника, когда он приблизится к ней на 2-3 м (рис. 12.5). Клетка А2 генерирует нервные импульсы только в ответ на громкие звуки. Она начинает реагировать, когда летучая мышь близко, и, вероятно, ее импульсы прерывают действие механизмов ЦНС, управляющих полетом. В результате он становится беспорядочным, и бабочка устремляется к земле. Наблюдения показывают, что с помощью такого уклоняющегося движения бабочки спасаются от летучих мышей как раз в тот момент, когда они приближаются на расстояние непосредственного нападения. Таким образом, ис-
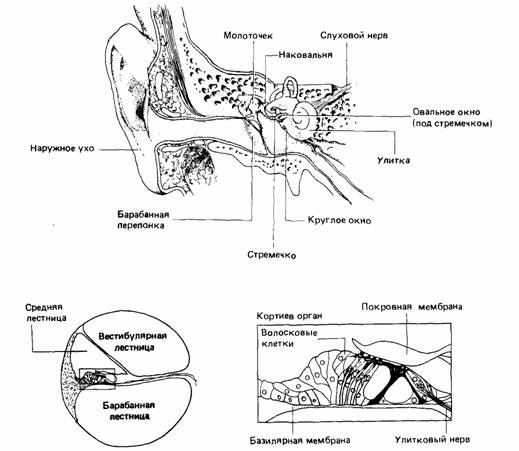
следования Редером (Roeder, 1963, 1970) слуха у бабочек служат прекрасной иллюстрацией не только механизмов работы простого уха, но и того, что сенсорный аппарат животного тонко адаптирован к его экологии. Звук обладает многими свойствами, на которые животное может реагировать. Когда он проходит через среду, ее частицы движутся взад и вперед, создавая осциллирующие волны давления. Величина (амплитуда) этих волн определяет интенсивность (громкость) воспринимаемого звука. Скорость его зависит от плотности среды распространения и не зависит от интенсивности. В воздухе звук распространяется со скоростью около 340 м в секунду, причем в горячем воздухе - быстрее, чем в холодном. В воде его скорость приблизительно в четыре раза выше, чем в воздухе. Если мы изобразим звук в форме волн, как на рис. 12.6, то расстояние между последовательными пиками, называемое периодом, обратно пропорционально частоте, т. е. числу пиков в единицу времени. Частота звука измеряется в герцах, или в циклах, в секунду. Простейший звук - чистый тон, единственная частота которого субъективно воспринимается как высота. Естественные звуки редко бывают чистыми тонами и состоят главным образом из ряда смешанных частот. Когда сложный звук разлагают на составляющие частоты, получают так называемый звуковой спектр. Орган слуха может быть чувствителен к широкому диапазону частот; например, тимпанальный орган саранчи реагирует на частоты от 1000 до 100 000 Гц. Когда слуховой рецептор воспринимает узкий частотный диапазон, говорят, что он «остро настроен». Антеннальный рецептор (джонстонов орган) самца комара Aedes aegypti реагирует на колебания частотой от 150 до 550 Гц, что соответствует звуку, создаваемому крылом самки. Высокочастотный тон от крыла самца не воспринимается джонстоновым органом (Haskell, 1961). В целом органы слуха позвоночных чувствительны к более широкой области звукового спектра, чем у беспозвоночных. Так, лишь очень немногие беспозвоночные (причем их существование еще строго не доказано) могут различать две частоты, если только не обладают двумя по-разному настроенными типами рецепторов (Haskell, 1961). Для позвоночных такое различение не составляет проблемы благодаря отчасти строению их уха, а отчасти анализирующей роли ЦНС. У человека (рис. 12.7), подобно большинству млекопитающих, ухо разделено на три отдела: наружное, среднее и внутреннее ухо. Наружное ухо состоит из ушной раковины и слухового прохода, который отделяется от среднего уха барабанной перепонкой. Среднее ухо образовано полостью, лежащей за барабанной перепонкой и соединенной с глоткой через евстахиеву трубу - проход, который делает возможным выравнивание воздушного давления в наружном и среднем ухе. Пассажиры самолета иногда испытывают боль в ушах при резком изменении высоты во время подъема или приземления из-за разницы давления по обе стороны барабанной перепонки. Прохождение воздуха через евстахиеву трубу устраняет эту разницу, чему способствуют жевание, глотание или зевота. В среднем ухе находятся три маленькие косточки: молоточек, наковальня и стремечко, которые соединяют барабанную перепонку с другой перепопкой- овальным окном в стенке камеры среднего уха. Внутреннее ухо представляет собой лабиринт взаимосвязанных наполненных жидкостью камер и каналов. Оно состоит из двух различных частей: вестибулярного аппарата, который ведает чувством равновесия, и улитки - спирально свернутой трубки, которая служит органом слуха. Внутри улитки проходят три канала: вестибулярная лестница, берущая начало у овального окна; барабанная лестница, которая начинается у круглого окна и другим своим концом соединяется с вестибулярной, и средняя лестница, расположенная между первыми двумя. Среднюю и барабанную лестницы разделяет основная (базилярная) мембрана, на которой находится кортиев орган. Этот орган образован рядами рецепторных клеток с сенсорными волосками на апикальных концах. Волоски упираются в студенистую покровную (текториальную) мембрану. Дендриты сенсорных нейронов заканчиваются на поверхности волосковых клеток, и когда колебания основной мембраны заставляют эти клетки колебаться, деформация волосков вызывает генераторные потенциалы, которые возбуждают сенсорные нервы. Колебания воздуха улавливаются ушной раковиной и проходят по наружному слуховому проходу, заставляя барабанную перепонку колебаться с той же частотой. Эти колебания передаются через полость среднего уха по трем слуховым косточкам, образующим систему рычагов, которая уменьшает амплитуду колебаний, но увеличивает их силу. Кроме того, колебания крупной барабанной перепонки передаются на овальное окно гораздо меньших размеров; в результате звуковое давление на барабанную пере-
понку усиливается на овальном окне приблизительно в 22 раза, что улучшает обнаружение слабых звуков. Движение мембраны овального окна вызывает соответствующее движение жидкости в улитке. При прогибании мембраны внутрь улитки жидкость проталкивается из вестибулярной лестницы в барабанную, отчего мембрана круглого окна выгибается наружу, и давление в улитке снижается. За полный цикл жидкость движется сначала в одну, а затем в другую сторону. Эти движения в улитке происходят с частотой колебаний наружного воздуха. Они вызывают бегущую волну в основной мембране, и, отклоняясь вверх и вниз, она деформирует волосковые клетки, упирающиеся в покровную мембрану. Эта деформация возбуждает сенсорные нейроны. Точка максимальной амплитуды колебаний мембраны в улитке меняется с частотой звукового стимула. Еще в 1867 г. Гельмгольц, исходя из анатомических соображений, правильно постулировал, что высокочастотные волны фокусируются вблизи основания улитки, а низкочастотные оказывают максимальное действие у ее вершины. Современными представлениями о работе улитки мы обязаны инженеру связи Бекеши (Bekesy, 1952, 1960), получившему за свои исследования Нобелевскую премию. Он наблюдал за процессами внутри улитки, удалив из нее жидкость и заменив ее суспензией угля и распыленного алюминия. По отражению
вспышек яркого света от этой суспензии он смог наблюдать прохождение волны в основной мембране. Как показали его наблюдения, эта мембрана натянута сильнее у основания, что благоприятствует высокочастотным колебаниям, и слабее у вершины, что благоприятствует низким частотам. Таким образом, определенные частоты колеблют различные участки основной мембраны и каждый участок стимулирует особые рецепторы кортиева органа. Синапсы нервных волокон, идущих от этих рецепторов, находятся в спиральном ганглии, а аксоны нейронов этого ядра образуют VIII черепномозговой нерв. Каждый из них сигнализирует об определенной частоте звука улитковому ядру головного мозга. Не у всех позвоночных строение уха одинаково. Так, например, у рыб и китообразных (дельфинов и китов) нет наружного уха, а рыбы лишены также барабанной перепонки и среднего уха со слуховыми косточками. Поскольку ткани рыб имеют приблизительно ту же плотность, что и вода, колебания, приходящие к их голове, могут передаваться прямо к внутреннему уху. Впрочем, некоторые рыбы обладают другим механизмом, функционально аналогичным среднему уху: это наполненный газом плавательный пузырь, у которого может быть костная связь с внутренним ухом, значительно улучшающая слуховую способность. У амфибий и рептилий самой наружной частью уха является барабанная перепонка, но у птиц уже имеется внешний канал (слуховой проход), ведущий к ней от поверхности тела. У птиц от внутренней поверхности барабанной перепонки идет костный стерженек (columella), соединяющийся со стремечком. У амфибий и рептилий эти косточки составляют часть челюсти, хотя у некоторых видов они играют определенную слуховую роль. Органы боковой линии у рыб и водных амфибий чувствительны к колебаниям, включая низкочастотные звуки; они состоят из видоизмененных волосковых сенсилл, которые реагируют на ток воды в канале боковой линии или на поверхности тела. Зрение Зрение основано на обнаружении электромагнитного излучения. Электромагнитный спектр имеет широкий диапазон, и видимая часть составляет лишь очень малую долю (рис. 12.8). Энергия электромагнитного излучения обратно пропорциональна длине волны. Длинные волны несут слишком мало энергии, чтобы активировать фотохимические реакции, лежащие в основе фоторецепции. Энергия коротких волн так велика, что они повреждают живую ткань. Большая часть коротковолнового излучения солнца поглощается озоновым слоем атмосферы: если бы этого не было, жизнь на Земле вряд ли могла возникнуть. Все фотобиологические реакции ограничены узким участком спектра между двумя этими областями. Фоторецепторные клетки содержат пигмент, который под действием света обесцвечивается. При этом изменяется форма молекул пигмента, причем в отличие от выцветания, с каким мы встречаемся в повседневной жизни, такой процесс обратим. Он ведет к еще не совсем понятным электрическим изменениям в рецепторной мембране (Prosser, 1973). Фоторецепторные клетки могут быть рассеяны по поверхности тела, как у дождевого червя (Lumbricus), однако обычно они образуют скопления. Глаз самого примитивного типа состоит из группы рецепторов, лежащих на дне углубления или ямки в коже. Такой глаз в общих чертах различает направление падаюшего света. Из-за теней, отбрасываемых стенками ямки, свет, падающий сбоку, освещает лишь одну ее часть, а остальная остается сравнительно темной. Такие различия в освещенности могут регистрироваться набором фоторецепторов в основании ямки. образующих зачаточную сетчатку. Глаз моллюска Nautilus с точечным отверстием (рис. 12.9) развился из глаза-ямки, внешние края которого сошлись к центру, а слой фоторецепторов образовал сетчатку. Такой глаз работает точно так же, как фотокамера с точечным отверстием: свет от каждой точки попадает только на очень малую область сетчатки, в результате возникает перевернутое изображение. Эволюцию глаза можно проследить у ныне живущих моллюсков, как показано на рис. 12.9. Из глаза Nautilus с точечным отверстием развился глаз с защитным слоем, вероятно, для предохранения от грязи. Внутри глаза образовался примитивный хрусталик, как у улитки Helix. Глаз такого типа обнаружен также у пауков. Встречаются и некоторые его разновидности, например глаз у гребешка Pecten, который имеет инвертированную сетчатку и зеркальную выстилку - тапетум (см. гл. 13). Глаз каракатицы Sepia (рис. 12.9) очень похож на глаз позвоночных. В нем находятся ресничные мышцы, которые могут менять форму хрусталика, и радужка, регулирующая, как диафрагма, количество падающего на сетчатку света. Глаза позвоночных, хорошим примером которых служит глаз человека, построены по единому плану, хотя, как будет показано в гл. 13, и у них отмечается некоторая экологическая адаптация. На рис. 12.10 показан горизонтальный разрез человеческого глаза. Он окружен плотной оболочкой - склерой, прозрачной в передней части глаза, где она называется роговицей. Непосредственно изнутри роговица покрыта черной выстилкой - сосудистой оболочкой, которая снижает пропускающую и отражающую способность боковых частей глаза. Сосудистая оболочка выстлана изнутри светочувствительной сетчаткой, которую мы более детально рассмотрим позднее. Спереди сосудистая оболочка и сетчатка отсутствуют. Здесь находится крупный хрусталик, делящий глаз на переднюю и заднюю камеры, заполненные соответственно водянистой влагой и стекловидным телом. Перед хрусталиком расположена радужка - мышечная диафрагма с отверстием, называемым зрачком. Радужка регулирует размеры зрачка и тем самым количество света, попадающее в глаз. Хрусталик окружен ресничной мышцей, которая изменяет его форму. При сокращении мышцы хрусталик становится более выпуклым, фокусируя на сетчатке изображение предметов, рассматриваемых вблизи. При расслаблении мышцы хрусталик уплощается и в фокус попадают более отдаленные предметы. У позвоночных в отличие от таких головоногих моллюсков, как каракатица, сетчатка имеет инвертированное, т. е. перевернутое, строение. Фоторецепторы лежат у сосудистой оболочки, и свет попадает на них, пройдя через слой нейронов главным образом ганглиозных и биполярных клеток. Ганглиозные клетки примыкают к стекловидному телу, и их аксоны проходят по внутренней поверхности сетчатки к слепому пятну, где они образуют зрительный нерв и выходят из глаза. Би-
полярные клетки - это нейроны, соединяющие ганглиозные клетки с фоторецепторами (рис. 12.11). Фоторецепторы делятся на два типа - палочки и колбочки. Палочки, более вытянутые по сравнению с колбочками, очень чувствительны к слабому освещению и обладают только одним типом фотопигмента - родопсином. Поэтому палочковое зрение бесцветное. Оно также отличается малой разрешающей способностью (остротой), поскольку много палочек соединено только с одной ганглиозной клеткой. То, что одно волокно зрительного нерва получает информацию от многих палочек, повышает чувствительность в ущерб остроте. Палочки преобладают у ночных видов, для которых важнее первое свойство. Колбочки наиболее чувствительны к сильному освещению и обеспечивают острое зрение, так как с каждой ганглиозной клеткой связано лишь небольшое их число. Они могут быть разных типов, обла-
дая специализированными фотопигментами, поглощающими свет в различных частях спектра. Таким образом, колбочки служат основой цветового зрения. Они наиболее чувствительны к тем длинам волн, которые сильнее всего поглощаются их фотопигментами. Зрение называют монохроматическим, если активен лишь один фотопигмент, например в сумерках у человека, когда работают только палочки (рис. 12.12). Дихроматическим зрение бывает при наличии двух активных фотопигментов, как у серой белки (Sciurus carolinensis) (рис. 12.12). Каждая длина волны стимулирует оба типа колбочек, но в разной степени в соответствии с их относительной чувствительностью в этой части спектра. Если мозг может распознавать такую разницу, животное различает длину волны света по его интенсивности. Однако эти определенные отношения возбудимости характерны более чем для одной части спектра, поэтому некоторые длины волн воспринимаются одинаково. Это происходит также при особых формах цветовой слепоты у человека. Длина волны, одинаково возбуждающая оба типа колбочек (в области пересечения кривых поглощения), воспринимается как белый цвет и называется «нейтральной точкой» спектра. Наличие ее показано в поведенческих опытах у серой белки (Muntz, 1981). Такое смешение меньше выражено в зрительных системах с тремя типами цветовых рецепторов или при трихроматическом зрении (рис. 12.12), известном у многих видов, в том числе у человека. Однако некоторое смешение происходит и здесь: можно, например, вызвать впечатление любого цвета посредством разных сочетаний трех монохроматических составляющих, специально подобранных по интенсивности и насыщенности. Без этого было бы невозможно зрительное восприятие цветной фотографии и цветного телевидения. У многих птиц и рептилий обнаружено больше трех типов цветовых рецепторов. Кроме различных фотопигментов, колбочки этих животных часто содержат окрашенные капельки масла, которые действуют как фильтры и в сочетании с фотопигментом определяют спектральную чувствительность рецептора (рис. 12.13). Эти капельки обычно не распределены по сетчатке равномерно, а сосредоточены в определенных ее частях. В 1825 г. чешский физиолог Ян Пуркинье заметил, что красные цвета кажутся ярче синих днем, но с наступлением сумерек их окраска блекнет раньше, чем у синих. Как показал в 1866 г. Шульц, это изменение спектральной чувствительности глаза, названное сдвигом Пуркинье, объясняется переходом от колбочкового
зрения к палочковому во время темновой адаптации. Это изменение чувствительности при темновой адаптации можно измерить у человека, определяя порог обнаружения едва видимого света через разные промежутки времени пребывания в темной комнате, По мере адаптации этот порог постепенно снижается, как показано на рис. 12.14. Перегиб кривой обусловлен переходом от колбочкового зрения к палочковому. Долю колбочкового зрения можно определить, направляя очень слабый свет на центральную ямку на сетчатке, в которой палочки отсутствуют. Долю участия в восприятии палочек определяют у «палочковых монохроматов», т. е. у редких индивидуумов, лишенных колбочек. Как можно видеть на рис. 12.14. палочки гораздо чувствительнее к свету, чем колбочки, но содержат только один фотопигмент - родопсин, максимальная чувствительность которого лежит в синей части спектра. Поэтому синие предметы кажутся в сумерках ярче предметов других цветов. Диапазон интенсивности света, воспринимаемого глазами позвоночных, огромен - они чувствительны к значениям освещенности, различающимся в миллиард раз. Это достигается разными механизмами, особыми для каждого вида. У многих рыб, амфибий, рептилий и птиц пигмент сосудистой оболочки концентрируется между наружными сегментами рецепто- ров при сильном освещении и оттягивается назад при его ослаблении. У этих животных наружные сегменты колбочек также подвижны. У некоторых рыб и амфибий в противоположном направлении движутся и наружные сегменты палочек. Количество света, достигающего сетчатки, регулируется сокращением зрачка. Этот рефлекс хорошо развит у угрей и камбал, ночных рептилий, птиц и млекопитающих (Prosser. 1973). Для того чтобы на сетчатке возникало резкое изображение, проходящий в глаз
свет должен преломляться так, чтобы фокусироваться на ней. Это происходит в роговице и хрусталике. В глазу человека преломление в роговице примерно вдвое больше, чем создаваемое хрусталиком. Трудность в том, что роговица отстоит от сетчатки на фиксированное расстояние, и поэтому для того, чтобы предметы, находящиеся на разных расстояниях, попадали в фокус, требуется некоторая аккомодация. Для этого предназначен хрусталик. У рыб он почти сферический, с высоким показателем преломления и малым фокусным расстоянием. Это необходимо потому, что показатель преломления воды почти такой же, как у роговицы, и на поверхности глаза преломления не возникает. Форма хрусталика не постоянна, и аккомодация достигается изменением расстояния между ним и сетчаткой. У наземных позвоночных аккомодацией управляют ресничные мышцы, которые изменяют форму хрусталика. При фокусировке близко лежащих предметов хрусталик становится более сферическим, а при фокусировке отдаленных объектов он уплощается (рис. 12.15). Животные, живущие как в воде, так и вне ее, не способны видеть хорошо в обеих средах. Глаза лягушки, крокодила и гиппопотама расположены в верхней части головы, и животное может видеть объекты над поверхностью воды, когда тело его погружено. У так называемых четырехглазковых рыб (Anableps anableps) каждый глаз разделен надвое полоской кожи. Верхняя его часть выступает над водой, когда нижняя остается погруженной. Единственный хрусталик в каждом глазу овальный и имеет такую форму, что нижняя часть может фокусировать подводные объекты, а верхняя - предметы над поверхностью воды. Поле зрения у глаз во многом зависит от их положения на голове. У позвоночных поле зрения каждого глаза составляет около 170° (Duke-Elder, 1958). У разных видов поля зрения обоих глаз перекрываются в разной степени. В общем, у хищников это перекрывание спереди значительное, а сзади
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 757; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.014 с.) |


 Рис. 12.3. Распределение и строение вкусовых почек человека.
Рис. 12.3. Распределение и строение вкусовых почек человека.