Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Чувствительные периоды в развитииСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Некоторые животные, по-видимому, запрограммированы на восприятие определенных аспектов среды в конкретные периоды своего развития, например освоение языка у человека. По сравнению с другими типами научения овладевание языком «запрограммировано» в том смысле, что оно происходит без каких-либо очевидных поощрений или наказаний. И еще пример. У птенцов многих выводковых видов птиц наблюдается совершенно неразборчивая привязанность к движущимся объектам: только что вылупившиеся утята кряквы, отсаженные от матери, будут следовать за грубой моделью утки, человеком или даже просто ящиком, медленно движущимися от них
(рис. 3.3). Для проявления этой реакции следования некоторые раздражители оказываются более эффективны, нежели другие (см. разд. 20.3). В естественной обстановке наиболее эффективным раздражителем является мать, и приближение к матери часто вознаграждается телесным контактом и теплом или пищей, которую мать находит. В лаборатории привязанность к модели может быть усилена пищевым подкреплением. Чем сильнее у животного развивается привязанность к одному объекту, тем меньше оно интересуется другими. Этот процесс научения, посредством которого обычно развивается привязанность к матери, называется импринтинг. Он имеет место в течение определенного чувствительного периода развития, продолжительность которого меняется в зависимости от вида и условий среды. Импринтинг может иметь долговременное влияние не только на привязанность к истинным или приемным родителям. У многих млекопитающих такой ранний опыт играет определенную роль в последующих социальных отношениях. Показано, что у ряда видов птиц импринтинг влияет на последующее социальное поведение (см. разд. 20.5). Короче говоря, импринтинг-это процесс научения, который имеет место на конкретных стадиях развития и который влияет на последующее поведение по отношению к родителям, собратьям или половым партнерам. Если объект привязанности представляет собой картонный ящик, то у утенка устанавливаются к этому ящику такие же отношения, как к родителю. Если самцы зебровой амади-ны (Taeniopygia guttata) выращиваются бронзовой амадиной (Lonchura striata), то, став взрослыми, они начинают ухаживать за самками бронзовой амадины.
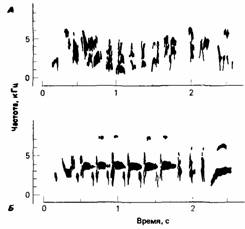
Хотя на этот тип научения, возможно, оказывает влияние поощрение, оно не зависит от поощрения или от каких-либо конкретных следствий поведения. Это научение запрограммировано как часть нормального процесса развития и при любых обстоятельствах свойственно определенному времени. Сходные механизмы функционируют при научении песне у некоторых воробьиных птиц. Первая попытка воспроизвести песню, называемую предпесней, обычно предпринимается молодыми птицами в первую весну или первую осень через несколько месяцев после вылупления. Предпесня напоминает взрослую песню по длине, высоте и тембру, но в ней отсутствуют некоторые элементы и украшения, типичные для взрослой песни; кроме того, предпесня обычно более изменчива и не столь точна (рис. 3.4). Если белоголовых воробьиных овсянок (Zonotrichia leucophrys) выращивать в изоляции, то у них развивается предпесня, но становления нормальной взрослой песни не происходит. У самцов белоголовой воробьиной овсянки, имеющих возможность слышать нормальную песню взрослого самца в возрасте от 10 до 90 сут, впоследствии
(т. е. в возрасте около 8 мес) развивается нормальная взрослая песня. Однако если они слышат эту песню только до 8-дневного возраста, то не могут впоследствии воспроизвести ее. Точно так же если самцы белоголовой воробьиной овсянки слышали эту песню после 100-дневного возраста, то освоить ее они уже не могут. Следовательно, существует критический период между возрастом 10 и 90 сут, когда молодым самцам белоголовой воробьиной овсянки необходимо слышать песню взрослого самца, для того чтобы они в конце концов научились петь эту песню (Marler, Mundinger, 1971). Такой же критический период характерен и для других видов. Если белоголовых воробьиных овсянок с помощью хирургических методов лишить слуха до того, как они услышат нормальную песню во время критического периода, нормальная песня у них не развивается. Если же их изолировать после прослушивания нормальной песни в критическом периоде, то тогда нормальная песня у них все же развивается. Таким образом, птицы, услышав эту песню во время критического периода, запоминают ее, хотя и не поют до 8-месячного возраста. Однако если их лишают слуха после прослушивания нормальной песни в критическом периоде, но до того, как они запоют, нормальная взрослая песня все равно не развивается (Konishi, 1965). Возможно, они слушают свое пение и используют слуховую память, чтобы развивать песню, соответствующую той, которую они слышали во время критического периода. Если белоголовых воробьиных овсянок лишить слуха после того, как песня отрепетирована и приобрела законченный вид, на песню это не повлияет. Следовательно, во время отработки и совершенствования песни она кодируется в другом виде памяти, возможно, как серия инструкций к пению. Поведение молодых животных Животное развивается на протяжении всей своей жизни и должно быть хорошо адаптировано к окружающей его среде всю жизнь. Таким образом, развитие определенных форм поведения - это не просто задача конструирования моделей взрослого поведения. У молодых животных часто наблюдается поведение, которое никогда более не проявится в их последующей жизни (рис. 3.5). Это поведение, как правило, предназначено для удовлетворения потребностей молодого животного. Например, состояние тревоги у птенцов серебристой чайки проявляется совершенно иначе, чем у взрослых животных. Испуганные птенцы отбегают недалеко от гнезда и затаиваются, молчащие и неподвижные, среди растительности. Взрослые же особи улетают прочь от гнезда, издавая крики тревоги. Птенцов, которые мечутся в испуге, легко могут схватить вороны или другие чайки. Оставаясь неподвижными, они усиливают возможности защитной окраски и уменьшают риск, что их заметит и съест хищник. Степень сходства поведения молодых и взросых животных различна у разных видов. У некоторых, например гну (Сопnochaetes), детеныши начинают следовать за стадом уже через несколько минут после рождения; в этом случае образ жизни молодых очень похож на образ жизни ихродителей. У других, например бабочек и лягушек, проходящих в своем развитии личиночные стадии, образ жизни молодых особей отличается от образа жизни взрослых. Жизненный цикл животного служит материалом для естественного отбора, поэтому для каждого вида характерна своя жизненная стратегия. И если влияние одних генов на поведение проявляется с самого начала развития, то действие других осуществляется с большой задержкой. Полагают, что такая поздняя экспрессия некоторых генов ответственна за процесс старения (Dawkins, 1976). Врожденное поведение Вопрос о том, какое поведение следует считать врожденным, долго был предметом споров. Термин «врожденный» имеет множество значений, которые необходимо различать. Ранние этологи считали, что врожденное поведение предопределено наследственностью, является неотъемлемой частью природы животного и потому не зависит от индивидуального опыта. Так, Лоренц (Lorenz, 1939) считает, что поведенческие признаки наследственны, закреплены индивидуально и, следовательно, их можно анализировать с эволюционных позиций. Точно так же Тинберген (1942) говорит об инстинктивных действиях как о чрезвычайно стереотипно скоординированных движениях, нейромотор-ный аппарат которых полностью включается в наследственную конституцию животного. Эти взгляды обоснованно критиковали Лерман (Lehrman, 1953) и другие исследователи, считавшие их слишком косными, включающими наивную идею генетического детерминизма и семантически запутанными. Строгое разграничение врожденного и приобретенного поведения не может удовлетворить этолога, поскольку на многие аспекты поведения влияют и генетические факторы, и индивидуальный опыт. Представление о том, что гены определяют поведение, наивно, так как в генах не может содержаться подробной информации для конкретных видов поведения. Гены могут влиять на процессы развития разными путями, но на эти же процессы влияют средовые факторы. Как мы уже видели, гены не могут предопределять течение онтогенеза независимо от окружающей среды, в которой происходит развитие. Для исследователей, изучающих поведение, удобно использовать термин врожденное поведение как сокращение для обозначения «поведения, которое развивается без очевидного влияния окружающей среды». В таком же смысле мы и будем употреблять его в этой книге.
ДЛЯ ЗАПОМИНАНИЯ 1. Развитие особи от зародыша до взрослого организма включает множество взаимодействий между генетическим аппаратом животного и его окружающей средой. Это взаимодействие предполагает, что каждая стадия развития служит основой для последующей. Этот процесс называется эпигенезом. 2. Влияние окружающей среды на развитие наиболее важно сразу же после рождения или вылупления, однако оно существенно и на других стадиях процесса развития. Условия, в которых родители выращивают детенышей, обычно предназначены для защиты детенышей от неблагоприятного влияния окружающей среды. 3. Для многих видов характерны периоды в развитии, когда особь чувствительна к определенным типам влияния извне. То, чему животное научается во время этого чувствительного периода, обычно сохраняется на протяжении всей его дальнейшей жизни. 4. Молодым животным часто свойственно поведение, которое позволяет им соответствующим образом реагировать на различные раздражители, поступающие из окружающей среды, такие, как появление хищника или родителей с пищей. Это типичное ювенильное поведение утрачивается во взрослом состоянии. 5. Для поведения, которое имеет место без очевидного влияния окружающей среды, удобно использовать термин «врожденный», не забывая при этом, что факторы окружающей среды в какой-то степени влияют на все поведение в целом. Рекомендуемая литература Bateson P.P.G., Klopfer H. (eds.) (1982). Perspectives in Ethology, v. 5, Ontogeny, Plenum, New York.
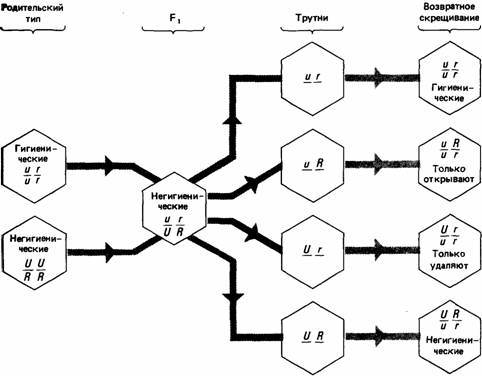
Генетика поведения Чарлз Дарвин, несомненно, интересовался, существует ли связь между наследственными факторами и поведением, однако пионером в этой области науки был другой внук Эразма Дарвина - Фрэнсис Гальтон. «Происхождение видов» побудило Гальтона посвятить остаток своей жизни изучению наследования психических особенностей. В 1869 г. Гальтон опубликовал книгу «Наследственная гениальность: исследование ее законов и значение». Он считал, что люди выдающихся умственных способностей чаще обнаруживаются среди родственников таких же выдающихся людей, чем среди населения вообще. Не имея какого-либо удовлетворительного метода, позволяющего оценивать умственные способности, Гальтон полагался на показатель репутации, «репутации лидера общественного мнения, изобретателя, человека, перед которым мир осознанно считает себя в большом долгу» (Gallon, 1869). Он проанализировал родословные почти 300 семей, в которых были выдающиеся судьи, государственные деятели, военачальники, писатели, ученые, поэты, музыканты, художники и т. д. Его результаты показали, что видного социального положения с наибольшей вероятностью достигают близкие родственники, а по мере того как родство становится дальше, вероятность стать выдающимся уменьшается. Гальтон сознавал, что, как правило, выдающиеся люди имеют одинаковые социальные и финансовые преимущества, а также возможности получить образование. Он считал, что репутация выдающихся людей указывает на их природные способности и не обусловлена средовыми факторами. Чтобы обосновать свои аргументы, Гальтон указывал на то, что многие выдающиеся люди произошли из семей ничем особенным не выделяющихся. Он пришел также к выводу, что приемные дети римских католических пап, которые пользуются огромными социальными благами, составляют группу менее известных людей, чем дети выдающихся деятелей. Гальтон затратил значительные усилия, чтобы улучшить способы оценки психических характеристик. Он разработал методики для оценки остроты обоняния, осязания и зрения и для оценки роста и веса, времени реакции и объема памяти. В статистике он ввел понятия корреляции, медианы и процентиля. В 1883 г. он ввел метод исследования близнецов для выяснения относительного влияния генотипа и среды. Он изучил 35 пар идентичных близнецов, которые были абсолютно схожи при рождении и воспитывались в одинаковых условиях. Данные его анализа показали, что их поведенческое сходство сохраняется, даже если во взрослом состоянии они живут раздельно. Галь-тоном были также проанализированы 20 пар неидентичных близнецов, которые были не столь похожи при рождении, но воспитывались в одинаковых условиях. Он писал: «Никуда не денешься от вывода, что природа чрезвычайно сильно преобладает над воспитанием, когда различия в воспитании не превышают того, что обычно имеет место среди людей одинакового социального положения в одной стране. У меня есть опасения, что мои данные, возможно, доказывают слишком многое и могут восприниматься с недоверием, потому что сильно противоречат житейскому опыту, согласно которому воспитанием можно добиться столь немногого» (Gallon, 1883). После работ Гальтона стали быстро развиваться исследования генетической основы поведенческих признаков, однако до сих пор в этих исследованиях содержится много противоречий. В начале нашего столетия наибольший интерес вызвал бихевиоризм. В частности, работы Уотсона (Watson, 1930) убедили многих психологов, что развитие поведения не определяется генетическими факторами, и твердые взгляды сторонников влияния средовых факторов на поведение, принятые бихевиористами, оставались господствующими вплоть до шестидесятых годов. Можно сказать, что как наука генетика поведения ведет свое начало с 1960 г., когда Фуллер и Томпсон (Fuller, Thompson, 1960) опубликовали книгу «Генетика поведения». В ней представлена история исследований поведения и умственных способностей человека и рассмотрены данные, свидетельствующие о влиянии генов на поведение. Несмотря на достаточную обоснованность данных, многие социологи и психологи остались противниками идеи влияния генетических факторов на поведение, и споры продолжались (см. Hirsch, 1963, 1967). Даже сегодня некоторые теории, основанные на генетической аргументации, остаются противоречивыми. В этой главе мы рассмотрим данные, свидетельствующие о влиянии генетических факторов на поведение. Единичные гены и поведение Поведенческие признаки, контролируемые отдельными генами, могут быть предметом строгого генетического анализа поведения. Классический пример этого-работа Маргарет Басток (Margaret Bastock, 1956), изучавшей условия успешного спаривания у плодовой мушки Drosophila melanogaster. Эта исследовательница скрещивала особей, несущих мутацию yellow (желтое тело), с дикими мухами в течение семи поколений. Полученные данные свидетельствовали о том, что дикие мухи генетически сходны с носителями мутации yellow, за исключением участка хромосомы, где локализован ген yellow. Басток обнаружила, что самцы, несущие мутацию yellow, реже спариваются с дикими самками, чем дикие самцы. У желтотелых самцов изменен ритуал ухаживания, и это снижает их успех при размножении. Они в меньшей степени стимулировали самок к спариванию, так как во время ухаживания слишком мало вибрировали крыльями. Ротенбюлер (Rothenbuhler, 1964) провел тонкий генетический анализ гнездоочистительного поведения медоносной пчелы (Apis mellifera). Личинки этих пчел иногда погибают от болезни, именуемой американской пчелиной гнильцой. Чтобы поддерживать гигиенические условия внутри улья, нормальные рабочие пчелы вскрывают ячейки, в которых находятся больные личинки, и удаляют их. Некоторые расы пчел, называемые «негигиеническими», этого не делают. Скрещенные с нормальными гигиеническими пчелами, они дают негигиенических потомков, что свидетельствует о доминантности этого признака. При скрещивании гибридов с родительской гигиенической расой Ротенбюлер получил следующие результаты. Из 29 образованных колоний в девяти зараженные ячейки оказались вскрытыми, но больные личинки из них удалены не были; в шести - ячейки не вскрывались, но если их вскрывал экспериментатор, то пчелы удаляли больных личинок; в восьми колониях пчелы не вскрывали ячейки и не удаляли личинок. На основе полученных данных Ротенбюлер предположил, что вскрывание ячеек и удаление личинок контролируется различными генами. Эти результаты можно объяснить, постулируя существование двух пар аллелей, из которых негигиенические аллели доминантны. Таким образом, рабочие пчелы с генотипом Uu или UU небудут вскрывать зараженные ячейки, а с Rr или RR не будут удалять личинок. У гаплоидных самцов пчел, которых называют «трутнями», только один набор хромосом. В 29 семьях Ротенбюлер обнаружил четыре типа трутней (UR, Ur, uR, ur), которых он скрестил с гигиеническими матками (uu, rr), как показано на рис. 4.1. В соответствии с этой простой менделевской схемой возвратное скрещивание дает четыре генотипа в равных соотношениях, и результаты, полу-
четные Ротенбюлером, существенно не отличаются от нее. Среди полностью гигиенических, частично гигиенических или полностью негигиенических рабочих пчел необнаружено ни физических, ни физиологических различий, хотя имеются данные, что негигиенические пчелы все же могут проявлять гигиеническую активность, но очень редко, и требуют для этого более мощных стимулов, чем нормальные. Это заставляет думать, что аллели U и u действуют как переключатели, которые запускают поведение вскрывания ячеек при условии, что существует некий порог стимуляции. Гены, которые действуют как переключатели, активируя группу других генов, известны и в ряде других случаев. Например, у некоторых бабочек весьма сложный рисунок крыльев напоминает рисунок крыльев других видов, несъедобных для хищников. Развитие мимикрического рисунка контролируется множественными генами, но оказывается, что определяет появление рисунка единственный ген-переключатель (Sheppard, 1961). Единичные гены известны также и у человека. Примером может служить недостаточность лак-тазы, обнаруженная у некоторых расовых групп (см. разд. 5.2). Синтез лактазы контролируется геном с тремя аллелями - L, l 1 и l 2. Как l 1, так и l 2 рецессивны по отношению к L, а l 2 рецессивен по отношению к l 1. У особей с генотипами LL, Ll 1 или Ll 2 лактаза синтезируется как у взрослых, так и у детей. Особи с генотипом l 1 l 1 или l 1 l 2 не синтезируют лактазу во взрослом состоянии, а особи с l 2 l 2 не могут ее синтезировать даже в детстве. Взрослые люди с генотипами l 1 l 1 или l 1 l 2 могут употреблять в пищу только кисломолочные продукты. Хромосомные мутации Расположение и число хромосом часто можно наблюдать непосредственно под микроскопом. Известны различные типы
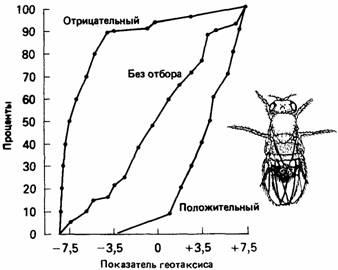
хромосомных мутации и влияние некоторых из них на фенотип. Поэтому изучение связи между поведением и строением хромосом представляет собой вполне подходящий метод для исследования влияний генов на поведение. Излюбленным объектом такого типа исследований является плодовая мушка Drosophila. В клетках слюнных желез личинок дрозофилы содержатся гигантские хромосомы; препараты слюнных желез сравнительно легко приготовить и анализировать. Впервые хромосомный анализ при изучении поведения дрозофилы применили Хирш (Hirsch) и его коллеги. Они изучали стремление Drosophila melanogas-ter двигаться в направлении силы земного притяжения или против него (положительный или отрицательный геотаксис). Поведенческую реакцию они проверяли в вертикальном пластиковом лабиринте (рис. 4.2), который должны были преодолеть мухи, привлекаемые запахом пищи. Обратному движению препятствовали конусообразные воронки в соединениях лабиринта. Большое число мух запускали в пробирку с левой стороны лабиринта и собирали в серию пробирок справа. Таким способом можно проверить тысячи мух, не трогая их при этом руками. Среди мух, собранных в различные пробирки с правой стороны, можно было выделить мух со строго положительным геотаксисом, со строго отрицательным геотаксисом и промежуточных. Скрещивая мух из крайней верхней и крайней нижней пробирок, можно получить расы с положительным или отрицательным геотаксисом (рис. 4.3). В одном из экспериментов сравнивали три популяции дрозофил (Hirsch, Erlen-meyer-Kimling, 1962). В одной вели отбор на положительный геотаксис, в другой - на отрицательный геотаксис, а в третьей, контрольной, популяции проводилось неселективное скрещивание. Мух скрещива-
ли с особой линией, в которой были различные хромосомные инверсии и гены-маркеры. У Drosophila melanogaster имеются четыре пары хромосом, три пары больших и одна - маленьких. Гены-маркеры использовались для идентификации трех больших хромосом. Это были доминантные гены, контролирующие признаки, отчетливо проявляющиеся в фенотипе. Специальная схема скрещивания позволила получить самок, которые были либо гомозиготны, либо гетерозиготны по хромосомам, подлежащим исследованию. Эти три хромосомы можно было идентифицировать с помощью генов-маркеров следующим образом: хромосома Х-ген узких глаз (Bar), хромосома II-ген волнистых крыльев (Cy) и хромосома III - ген, обусловливающий короткие щетинки (Sb). Тестируемую самку, несущую эти маркеры, скрещивали с самцами одной из исследуемых линий. Для использования в последующих экспериментах из потомства отбирали только тех, которые несли все три гена-маркера. При возвратных скрещиваниях, используя самцов из исходной популяции, получили восемь возможных генотипов. Следовательно, каждая из трех больших хромосом гетерозиготна или гомозиготна по S-хромосоме, полученной от тестируемой линии (s), к которой принадлежал отец. На генотипах этих восьми классов изучали индивидуальные эффекты и взаимодействие хромосом. Хирш и Линда Эрленмейер-Кимлинг (Hirsch, Erlenmeyer-Kimling, 1962) обнаружили, что в популяции, где отбор не проводился, хромосомы X и II оказывают большое влияние на развитие положительного геотаксиса, а хромосома III-на развитие отрицательного геотаксиса. В линии, где проводился отбор на положительный геотаксис, были обнаружены небольшие изменения в хромосомах X и II, но хромосома III в этом случае оказывала влияние на развитие положительного геотаксиса. В линии, которая была отобрана на отрицательный геотаксис, отрицательное влияние хромосомы III увеличивалось, тогда как положительное действие хромосом X и II было меньше. Эффект от совместного действия всех трех хромосом был больше в случае отрицательного геотаксиса. Это неудивительно, так как общая реакция на отбор (см. рис. 4.3) сильнее в случае отрицательного геотаксиса. Эти результаты показывают, что поведение, связанное с геотаксисом, контролируется рядом генов, которые находятся во всех трех больших хромосомах. Хромосомный анализ широко применяется при изучении различных типов поведения дрозофилы, в том числе скорости спаривания и других аспектов ухаживания
(Ehrman, Parsons, 1976). Хромосомные инверсии обычны, и обнаружено, что D. pseudoobscura с такими инверсиями в гетерозиготном состоянии характеризуются большей приспособленностью, чем с ин-.версиями в гомозиготном состоянии, благодаря влиянию на поведение ухаживания. Хромосомные аномалии у человека являются предметом интенсивных исследований, и считают, что эти аномалии являются причиной таких заболеваний, как эпилепсия, маниакально-депрессивный психоз, умственная отсталость и шизофрения (McClearn, DeFries, 1973; Ehrman, Parsons, 1976).
|
||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 667; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.41 (0.02 с.) |