Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Причина возникновения смещенной активностиСодержание книги
Поиск на нашем сайте
Случаи смещенной активности были описаны еще ранними этологами (Huxley, 1914; Makkink, 1936; Kortlandt, 1940; Tinbergen, 1940). Впервые этот термин использовали Эдвард Армстронг (Armstrong, 1947), а также Нико Тинберген и Ян ван Йерсел (Tinbergen, van Iersel, 1947). Тинберген (1952) отмечает следующие особенности смещенной активности: «По своему внешнему проявлению смещенная активность напоминает видоспецифические комплексы действий животного или кажется производной от них». «Эти движения представляются абсолютно неуместными, поскольку они полностью выходят из контекста поведения, которое предшествует им или следует за ними». «Смещенная активность, по-видимому, проявляется в том случае, когда активированный драйв (специфическая для данного действия энергия) почему-либо не может разрядиться в виде соответствующего консумматорного акта (актов)».
Рис. 21.1. Самец трехиглой колюшки вентилирует свое гнездо. На рис. 21.2 показано несколько классических примеров смещенной активности. Кортланд (Kortland, 1940) и Тинберген (Tinbergen, 1940) высказали предположение о природе смещенной активности, которое вытекает из представления о том, что каждый инстинкт обладает своей собственной энергией, специфической для данного инстинктивного действия ("action-specific energy"). Кортланд различает автохтонное поведение, которое использует свою специфическую энергию (drive), и аллохтонное - использующее энергию, предназначенную для других действий. С этой точки зрения смещенная активность рассматривалась как аллохтонная. Тинберген (Tinbergen, 1952) считает, что «смещенная активность - это выходное отверстие, через которое может выразить себя в действии чем-то сдерживаемая специфическая энергия (the thwarted drive). Эта концепция обозначалась словом Ubersprung - перескок, или sparking-over - перевоспламенение (переброска пламени). Тинберген (Tinbergen, 1951) отмечал, что смещенная активность проявляется в том случае, когда «налицо избыточная мотивация, а нормальные пути для ее удовлетворения каким-то образом заблокированы». Он отождествлял причину такого блока с конфликтом двух предельно активированных антагонистических энергий или с высокой мотивацией, которая возникает в ситуации, где отсутствуют внешние стимулы, необходимые для реализации соответствующего консумматорного поведения. Конфликт между двумя источниками антагонистических энергий можно увидеть в ситуации, когда животные находятся на границе своих территориальных участков и защищают их. Так, например, самцы колюшки, встретившись на границе своих территорий, сразу принимают позы с опущенными вниз головами (рис. 21.3). Тинберген (Tinbergen, 1951) отождествляет эту позу с позой копательного поведения, которым начинается строительство гнезда: «Удивительно, что смещенная активность часто проявляется в таких ситуациях, когда специфические энергии защитного поведения и поведения избегания бывают активированы одновременно. В пределах своей территории самец неизменно атакует любого чужого самца. За ее пределами тот же самец не нападает на чужака, а напротив, уплывает от него. В промежуточной ситуации, т.е. на границе двух территорий, самцы-хозяева выполняют смещенные действия. Напрашивающийся отсюда естественный вывод о том, что смещенная активность в этой ситуации представляет собой «выходное отверстие» для конфликтующих энергий атаки и бегства (которые, естественно, одновременно не могут разрядиться самостоятельно, поскольку их моторные комплексы антагонистичны), был проверен в эксперименте (Tinbergen, 1940). Самцу колюшки на его территории показывали красную модель, на которую, конечно, он тут же нападал. Эксперимент был спланирован таким образом, что модель не «обращалась в бегство», а «оказывала сопротивление», ударяя атакующего самца. Если эта «контратака» выполнялась достаточно энергично, самец мог потерпеть поражение на своей собственной территории. Он уплывал от модели и прятался среди растений. Если после этого модель неподвижно находилась на территории самца, то постепенно она стимулировала у него специфическую энергию защиты. Однако тенденция к бегству со временем уменьшалась. Постепенно бойцовская энергия пересиливала тенденцию к бегству, и через несколько минут самец уже снова начинал атаковывать модель. Но непосредственно перед атакой самец выполнял смещенные копательные действия. Таким образом, этот эксперимент показывал, что смещенное копание происходит именно тогда,
Рис. 21.2. Примеры смещенной активности. 1 - смещенное гнездовое поведение во время агрессивных взаимодействий у серебристой чайки; 2 - смещенная реакция выдергивания травы (гнездо-строительное поведение) во время территориального спора; 3 — смещенная активность в виде сна во время драки между устричниками; 4 - то же у шилоклювок; 5 - смещенная реакция копания песка (гнездостроительное поведение) у самца трехиглой колюшки во время территориального спора; 6 смещенная реакция чистки перьев у пеганки во время ухаживания; 7 — та же реакция у чирка-трескунка; 8 - то же у мандаринки; 9 - то же у кряквы; 10 - то же у шилоклювки; 11 - смещенная реакция схватывания корма во время ухаживания у европейской голубой цапли; 12,13 и 14 - смещенное половое поведение европейского баклана во время агрессивных взаимодействий; 15 - смещенное выпрашивание пищи во время ухаживания у серебристой чайки; 16 - смещенное пищевое поведение во время драки у домашних петухов. (Из Tinbergen, 1951.)
когда две смещенные энергии описываемых действий достигают полного равновесия. Представляется весьма вероятным, что именно так следует объяснить природу смещенной активности, проявляющейся у самца во время защиты собственной территории». В настоящее время достаточно убедительно показано, что смещенная активность проявляется обычно в точке равновесия между двумя различными мотивационными тенденциями. Заметим, что само по себе это не является доказательством возникновения смещенной активности при избыточной мотивационной энергии. Однако, как мы увидим дальше, существуют и другие объяснения. При обзорном анализе полевых исследований (например, Tinbergen, 1952; Ваstock et al., 1953) было обнаружено, что смещенная активность проявляется, как правило, в трех различных ситуациях: 1) при физическом сдерживании аппетитив-
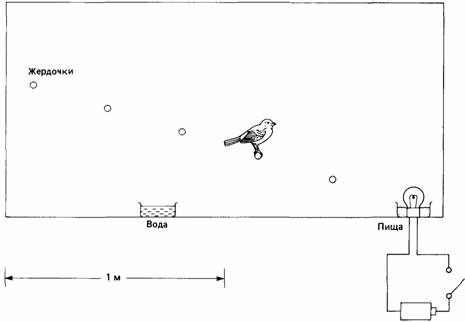
Рис. 21.3. Поза угрозы у самца трехиглой колюшки, которая сформировалась на основе реакции копания песка, предшествующей постройке гнезда. ного (подготовительного) поведения; 2) при сдерживании консумматорного (завершающего) поведения в результате удаления его объекта или цели и 3) при одновременной активации несовместимых тенденций. Эти наблюдения были подтверждены и в лабораторных исследованиях (например, McFarland, 1965). Общим для всех этих ситуаций представляется тот факт, что текущее поведение не может проявиться. Оно сдерживается либо физической преградой, либо недоступностью ожидаемого результата поведения, либо несовместимой активностью. В соответствии со своим классическим объяснением природы смещенной активности этологи считают, что эта активность аллохтонна, т.е. что она возникает не под действием своей специфической энергии, а в результате «разлива», или «перевоспламенения» энергии, сдерживаемой какими-либо причинами. Тинберген (Tinbergen, 1951) подразумевал также, что смещенная активность может облегчать напряжение и действовать как отдушина для выхода нервной энергии. По-видимому, такая точка зрения основана на интуитивных представлениях. Все мы неоднократно переживали субъективные ощущения напряжения, возникающие в определенных ситуациях, и можем предположить, что нервное манипулирование сигаретами или какими-либо висячими украшениями ослабляет это напряжение. Тинберген (Tinbergen, 1951) относил такой тип человеческого поведения к смещенной активности: «Другое явление, которое подразумевает наличие инстинктивной организации у человека, в основе своей напоминающей аналогичную организацию животных, - это смещенная активность. И у человека это совсем не редкое явление. Ее не так легко распознать, как у животных, поскольку у человека такие приобретенные моторные комплексы, как, например, зажигание сигареты, перебирание ключей, манипулирование носовым платком и т. д., зачастую носят характер смещенной активности. Вместе с тем у человека в качестве отдушины могут выступать и врожденные комплексы. Нередко наблюдающееся у человека почесывание за ухом в конфликтных ситуациях почти наверняка имеет врожденную основу. Удивительно, как часто в конфликтных ситуациях проявляются действия, относящиеся к инстинкту комфорта (уход за кожей); женщины главным образом начинают приводить к порядок прически, хотя этого и не требуется, а мужчины - бороды или усы; и делалось это не только в те дни, когда мужчины их носили, но проявляется и теперь - в нашу «чисто выбритую эпоху». Ранние этологи гордились тем, что они проводят объективное исследование поведения (например, Tinbergen, 1942), и конечно жаль, что они позволяли себе прегрешения субъективно спекулировать по поводу смещенной активности. Как мы увидим дальше, эта ситуация совсем не так проста, как они думали. Первые сомнения по поводу классической интерпретации смещенной активности возникли после того, как было обнаружено, что ее нельзя считать полностью аллохтонной. Так, например, индюки в самом разгаре боя могут вдруг внезапно прекратить его и начать демонстрацию смещенных действий: поедать пищу или пить воду (Raber, 1948). Какая именно смещенная активность проявится, зависит от того, что доступно птицам в данный момент: пища или вода. Эндрю (Andrew, 1956b) предположил, что смещенное поведение чистки перьев у вьюрков регулируется периферическими стимулами, которые в норме вызывают адекватные действия по чистке оперения. Он показал, что чистка перьев легко подавляется другими видами активности, и это дало ему основание утверждать, что стимулы, вызывающие у птиц чистку перьев, постоянно поступают из внешней среды. Эндрю полагал, что в конфликтных ситуациях, когда полностью не может проявиться ни один из конкурирующих видов активности, давление на поведение чистки перьев ослабевает до такой степени, что появляется возможность этому поведению проявиться в виде смещенной активности. В основу этой теории растормаживания, объясняющей природу смещенной активности, были положены данные, полученные главным образом при изучении поведения гнездящихся крачек (van Iersel, Bol, 1958). Когда у птиц возникал конфликт между стремлением остаться в гнезде и стремлением к бегству, они часто начинали «предаваться наслаждению» - смещенным действиям по чистке оперения. Ван Йерсел и Бол обнаружили, что такие смещенные действия продолжались более длительное время после дождя, т. е. именно тогда, когда в норме влажное оперение должно было бы служить стимулом для адекватных действий. Авторы пришли к заключению, что равновесие между двумя конфликтующими специфическими энергиями снижает их тормозящее воздействие на третий энергетический очаг, который поэтому получает возможность проявиться в форме открытого поведения. Подобным образом результаты по изучению смещенной активности - вентилирования гнезда у колюшки - показали, что эта активность проявляется именно тогда, когда оказываются в равновесии агрессивные и половые стремления (Sevenster, 1961). Смещенную активность можно было вызвать гораздо легче, если имитировали наличие в гнезде развивающихся икринок путем орошения гнезда водой, в которой была растворена двуокись углерода. Роуэлл (Rowell, 1961) исследовал на зябликах конфликтную ситуацию, возникающую при выборе этими птицами поведения приближения или поведения избегания. Такая ситуация создавалась двумя способами. В первом случае он показывал птицам чучело совы, к которому с криком приближались птицы, а во втором - пугал голодных птиц тем, что подавал внутри их кормушек вспышки света. В клетках, где проводились эти эксперименты, находились ряды жердочек, и экспериментатор мог определить, на какой именно жердочке стремления птицы двигаться вперед или, напротив, отступать находились в равновесии (рис. 21.4). Роуэллу удалось показать, что смещенное поведение чистки перьев и осушения клюва проявляется именно в точке этого равновесия и что на такие смещенные действия оказывают регулирующее влияние нормальные причинные факторы. Так, например, смещенное поведение чистки перьев наиболее отчетливо проявлялось именно тогда, когда птицу обливали смесью сажи и воды. По мнению Роуэлла, для возникновения смещенной активности необходимо, чтобы выполнялись три условия: 1) должно возникнуть состояние равновесия между двумя конкурирующими стремлениями животного или соответственно между двумя конкурирующими очагами специфической энергии; 2) такое равновесие должно продолжаться достаточно долго, чтобы смещенная активность могла проявиться; 3) необходимо наличие адекватных внешних раздражителей, вызывающих выполнение нормальных (а не смещенных) действий. Позднее было показано (McFarland, 1965), что наличие мотивационного конфликта не является непременным условием возникновения смещенной активности. В опытах на египетских горлицах, которым не давали воды, автор обнаружил, что в ситуациях выбора между стремлениями приблизиться к чему-либо или отступить у них проявляется смещенная активность, выражающаяся в виде клевания и чистки перьев, аналогичная той, что наблюдал Роуэлл. Вместе с тем смещенная активность проявлялась у птиц и тогда, когда на привычном пути к поилке ставилась стеклянная преграда или когда испытывающие жажду птицы, подойдя к поилке, не находили в ней воды. Смещенное клевание усиливалось, если птицы испытывали небольшой голод или на полу клетки обнаруживали зерна. Более того, горлицы, которые были нау-
Рис. 21.4. Экспериментальная установка для создания конфликта «приближение - избегание» у зяблика. Когда птица приближается к пище, перепрыгивая с жердочки на жердочку, экспериментатор включает находящуюся в кормушке лампочку, что вызывает у зяблика слабую тенденцию к избеганию. чены клевать освещенный диск, чтобы получить пищевое подкрепление, в ситуациях, где они испытывали жажду, но им препятствовали получить воду, выполняли это заученное пищедобывательное поведение как смещенную активность. Таким образом, по своей природе смещенная активность не всегда бывает врожденной. Эта первоначальная теория растормаживания, которую разрабатывали Эндрю (Andrew, 1956b), ван Йерсел и Бол (van Iersel, Bol, 1958), Роуэлл (Rowell, 1961) и Севенстер (Sevenster, 1961), согласуется с классической теорией избыточности специфической энергии в том пункте, где утверждается, что в конфликтных ситуациях смещенная активность проявляется в точках равновесия конфликтующих начал. Вместе с тем эта теория радикально отличается от классической тем, что признает необходимость мотивации для смещенной активности. Вместо того чтобы быть разрядкой избыточной энергии от другой активности, смещенная активность возникает в ответ на свои собственные стимулы, а это означает, что она автохтонна, а не аллохтонна. Значение мотивационного конфликта сводилось к достижению коротких периодов равновесия, или мотивационному тупику, во время которых и происходило растормаживание смещенной активности. Таким образом, последняя считалась активностью низкоприоритетной, которую можно было наблюдать лишь через «проломы», создаваемые в результате конфликтов между главными конкурирующими энергиями. Эту раннюю версию теории растормаживания критиковали (McFarland, 1966b) за то, что она определялась в терминах специфических энергий, или нервных центров, была нечетко сформулирована и признавала смещенную активность только в случаях мотивационного конфликта. Маргарет Басток и др. (Bastock et al., 1953) отмечали, что смещенная активность может проявиться как в ситуации конфликта, так и в случае, когда что-либо пре-
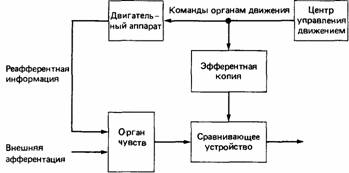
Рис. 21.5. Схема реафферентной системы. пятствует проявлению нормальной активности. Эти авторы высказали также предположение, что общим фактором во всех ситуациях, где наблюдается смещенная активность, может быть отсутствие определенного рода отрицательной обратной связи.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 764; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.167.85 (0.01 с.) |