Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Бюджет времени и бюджет энергииСодержание книги
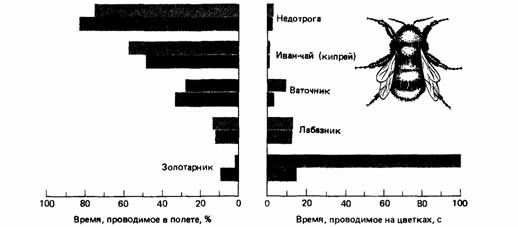
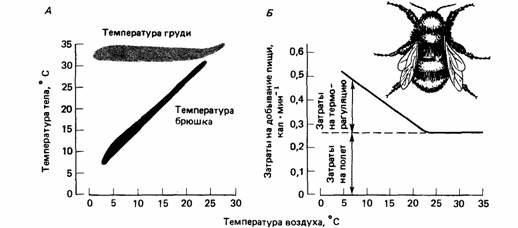
Поиск на нашем сайте Если на приобретение различных товаров и на осуществление какой-то деятельности мы располагаем ограниченной суммой денег, то мы часто распределяем эту сумму в соответствии с нашими нуждами. Обычно мы учитываем все наши расходы на какой-то определенный период времени - день, неделю или год - и называем это бюджетом. Можно составить бюджет предстоящих расходов, а можно - тех, которые уже были произведены. В одном случае мы ассигнуем определенные суммы денег на определенные цели, а в другом - мы подводим итог расходам, которые уже произвели. В любом случае ведение бюджета предполагает определенную дисциплину в расходовании денег. Мы уже видели, что деньги в потребительской экономике - это аналог энергии в поведении животных, поэтому естественно спросить, а есть ли у животных энергетический бюджет. Однако мы видели также, что энергия - это только один тип ограничителя, который воздействует на поведение животного. Другим важным ограничителем является время, и мы также можем задаться вопросом, приложимо ли понятие «бюджет времени» к поведению животных. Бюджет времени или энергии - это не просто подсчет того, как животное тратит время и энергию. Если бы животное растрачивало время и энергию совершенно беспорядочно, то это означало бы, что оно не имеет никакого бюджета. Однако мы можем ожидать, что естественный отбор сформировал животных такими, что они должны тратить имеющиеся у них энергию и время с максимальной пользой. Поэтому разумно ожидать, что животные используют время и энергию как ценные ресурсы и, следовательно, имеют соответствующие бюджеты. Хейнрих (Heinrich, 1979) сравнивал пищедобывательную деятельность шмеля (Bombus) с поведением покупателя: «Задача, с которой сталкивается шмель, начиная поиск пищи на лугу, где много различных растений, нисколько не отличается от задачи, с которой сталкивается неопытный покупатель, толкающий тележку вдоль торгового зала универсама. Прямо или косвенно они оба стараются получить за свои деньги как можно больше ценностей. Никто из них наперед не знает, что именно содержится в упаковках, которые стоят на полках или находятся на лугу. Но они научаются этому на опыте». Цветки обеспечивают шмелей энергией, необходимой для выращивания потомства, при этом шмели должны тратить значительное количество энергии на добывание пищи. Шмели способны жить в холодных климатических условиях благодаря их удивительной способности к терморегуляции. Они могут поддерживать высокую температуру тела при низкой температуре окружающего воздуха, что дает им воз- можность оставаться активными, даже если для этого потребуются высокие энергетические затраты. Они могут сберегать энергию, значительно уменьшая уровень активности и таким образом сохраняя тепло. Когда пищевые ресурсы цветков оказываются бедными, шмелям тем не менее удается извлекать выгоду за счет медленной пищедобывательной деятельности. Когда пищи очень много, шмели повышают температуру тела и интенсифицируют пищедобывательную активность. Следовательно, они рассчитывают свои энергозатраты в соответствии с имеющимися условиями. Собирая пищу, шмель проводит большую часть времени в полете. Перелетая с цветка на цветок, шмели стараются свести до минимума продолжительность и дальность полета. За час они пролетают от 11 до 20 км и проводят внутри гнезда всего 2-4 мин в паузах между полетами за пищей. Простые расчеты (Heinrich, 1979) показывают, что для добывающего пищу шмеля бюджет времени более важен, чем бюджет энергии. Предположим, например, что у одного шмеля цветки находятся близко от гнезда и он может постоянно «пастись» на них, тогда как другой шмель летает за взятком за три километра от гнезда. Если второй шмель пролетает по 15 км в час, то он должен тратить на каждый полет 24 мин. Питаясь на кипрее, оба шмеля смогут собрать полный зобик нектара (около 30 мг сахара) примерно за 10 мин. Шмель, который совершает дальние путешествия, добывает 30 мг сахара за 34 мин, но при этом около 3 мг сахара расходуется во время каждого полета на метаболические процессы. Таким образом, полеты туда и обратно отнимают у этого шмеля примерно две трети времени и только одну десятую часть энергии. За это же самое время шмель, питающийся вблизи гнезда, получил бы 102 мг сахара. Таким образом, имеет смысл тратить время на дальние перелеты лишь в том случае, если растения, растущие далеко от гнезда, в 3,4 раза ценнее, чем растения, растущие поблизости. Пищедобывательное поведение шмелей изменяется в зависимости от количества нектара. Чем больше нектара в каждом цветке, тем больше они обследуют других цветков в ближайших окрестностях. Хейнрих провел эксперименты, в которых закрывал тонкой прозрачной тканью некоторые участки клеверного поля, а другие участки оставлял открытыми. Шмели выбирали нектар из закрытого клевера, а в прикрытых цветках нектар накапливался. Когда ткань с цветков снимали и шмели могли посещать и богатые, и бедные нектаром участки клеверного поля, оказалось, что на богатом нектаром участке клеверника шмели обследовали около 12 цветков в головке клевера и совершали короткие перелеты между головками. На участке клеверника с низким содержанием нектара шмели проверяли примерно два цветочка в каждой головке и совершали более длинные перелеты между головками клевера (рис. 24.22). Именно таким путем шмели концентрировали свою пищедобывательную активность на самых выгодных участках клеверника. Энергетические затраты на добывание пищи в некоторой степени зависят от температуры окружающей среды. Когда шмели «пасутся» на кипрее, они задерживаются на каждом цветке всего лишь 1-2 с, собирая язычком крошечные капли нектара. Однако посещение других цветков длится до нескольких минут (рис. 24.23). Во время пребывания на цветке шмели не дают охладиться своему летательному аппарату и сохраняют его «полетную» температуру, что обеспечивает им готовность взлететь без всякой задержки. Во время длительного пребывания на цветке они поддерживают температуру грудной части тела около 32° С за счет дрожания грудных мышц; однако шмели не тратят энергии на согревание брюшка, которое не входит в состав двигательного аппарата, обеспечивающего полет (рис. 24.24). При температуре окружающей среды около 25°С шмелю нет необходимости согревать грудной отдел во время периодов покоя; если же температура падает ниже этой величины, то шмелю приходится «платить все дороже» для того, чтобы эффективно добывать пищу (рис. 24.24). В цветках разных видов растений образуется различное количество нектара, поэтому можно рассчитать относительные затраты на добывание пищи на
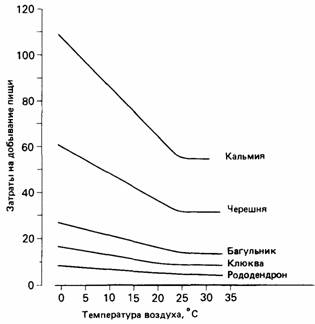
различных растениях при разной температуре воздуха (рис. 24.25). Шмели собирают пищу на «прибыльных» цветках рододендрона при самых различных температурах воздуха, тогда как на кальмии и черешне при низкой температуре они «пасутся» не столь интенсивно, поскольку эти растения не могут возместить им дополнительную энергию, требующуюся для терморегуляции (Heinrich, 1979). На примере пищедобывательной активности шмелей видно, каким образом время и энергия ограничивают ее эффективность. Когда шмелю приходится путешествовать на некоторое расстояние, чтобы найти медоносные растения, время становится лимитирующим фактором, и для шмеля выгодно потратить энергию, чтобы сберечь время. Когда же шмель питается на относительно бедных нектаром цветках или добывает пищу при низкой температуре, он может потратить больше времени ради того, чтобы сберечь энергию. Взаимоотношения между временем и
энергией часто играют важную роль в поведении животного. Ему приходится распределять эти ресурсы для каждого конкретного типа активности и для всех конкретных условий, с которыми оно сталкивается в данном случае. Кроме того, ему приходится принимать в расчет затраты на переход от одного типа активности к другому. Когда животное переключается с одного поведения на другое, это может потребовать определенных затрат. Например, питающийся на кукурузном поле голубь может захотеть пить, поскольку в данном случае пищу его составляют сухие зерна, а чтобы получить воду, ему необходимо пролететь более километра. Помимо физиологических затрат на перелет от места кормежки до места, где находится вода, птица будет тратить время на этот перелет и при этом не будет получать никакого выигрыша ни от приема пищи, ни от питья. Более того, во время этого перелета голубь может подвергнуться определенному риску при встрече с хищными птицами или вооруженными фермерами. Таким образом, затраты на переключение с одного вида активности на другую представляют собой уменьшение приспособленности, которое происходит в тот период времени, когда животное переходит от одной активности к другой и не получает пользы ни от одной из них. Эти затраты могут включать в себя потери ценного времени, расход энергии или риск от встречи с хищниками. Теоретически можно показать, что затраты на переключение следует рассчитывать таким образом, как будто они составляют часть затрат на активность, которую собирается осуществить животное. Следует различать мгновенные затраты, которые возникают на каждом отрезке соответствующего периода времени, и совокупные затраты за весь рассматриваемый период в целом (рис. 24.26). Весьма важным представляется то, что переключения с одного типа поведения на другое имеют смысл в том случае, если совокупные затраты вскоре после этого переключения станут меньше, чем они были бы, если бы животное вовсе не меняло своего поведения (рис. 24.26). Мною было экспериментально показано, что если затраты на смену активности высоки, то голуби переходят от пищевого поведения к питьевому менее часто, чем они делали бы это в других условиях. Вместе со Стефаном Ларкином (Larkin) мы получили данные о том, что птицы действительно включают затраты на смену активностей в состав затрат на
то поведение, которое они собираются осуществить (Larkin, McFarland, 1978). Они не сменят своей активности до тех пор, пока не рассчитают этих затрат. Ларкин (Larkin, 1981) обнаружил, что временная картина пищевого и питьевого поведения у голубей изменялась вполне предсказуемым образом, если затраты на переключение с одного вида поведения на другой увеличивались, т. е. возрастал расход времени или энергии. Помимо ежеминутного расчета затрат животные производят и более глобальную оценку бюджета своего времени и своей энергии. В целом можно ожидать, что поведение, основное назначение которого способствовать выживанию особи, будет иметь приоритет перед поведением, которое обеспечивает другие виды приспособленности, таким, например, как территориальное, половое или родительское. Однако виды животных значительно различаются между собой. Некоторые типы поведения существенны для животного всегда, тогда как другие (к примеру, терморегуляционное поведение) имеют значение только в тех случаях, когда исчерпываются возможности регуляторных физиологических механизмов. Так, для животных некоторых видов питьевое поведение - это каждодневная необходимость, тогда как животные других видов могут обходиться вовсе без него. Каждый вид активности имеет какое-то значение с точки зрения приспособленности животного, и животным приходится распределять по приоритету различные виды активности как в целом, так и в своей ежеминутной деятельности. Вместе с Хаустоном мы попытались решить эту проблему. Для этого мы оценивали, каковы будут затраты животного, если оно воздерживается от каждого вида активности, входящего в его естественный поведенческий репертуар (Houston, McFarland, 1980). Например, если животное не питается, его затраты будут высокими, однако если оно воздерживается от поведения чистки шерсти (grooming), затраты могут быть относительно низкими. Животное, испытывающее пищевую мотивацию и сильную мотивацию чистки шерсти, но имеющее слишком мало времени для удовлетворения обеих этих мотиваций, понесет меньшие потери с точки зрения своей общей приспособленности, потратив имеющееся у него время на питание. Предположим, что в течение обычного
дня своей жизни животное осуществляет различные виды полезного поведения таким образом, как это показано на рис. 24.27. Если условия, в которых живет животное, день ото дня практически не меняются, оно распределяет разные виды своего поведения в соответствии с имеющимся у него временем. А теперь представим себе, что внешняя среда изменилась таким образом, что животному требуется гораздо больше времени, чтобы получить обычное количество пищи (рис. 24.28). Оно может прореагировать на изменившиеся условия жизни по-разному: может тратить на питание то же самое время, что и раньше, но довольствоваться меньшим количеством пищи; будет стремиться получить обычное количество пищи; выбирать компромиссный вариант между этими двумя крайностями. Если животное будет тратить много времени на получение обычного количества пищи, то у него останется меньше времени на все другие виды активности, которые надо будет «сжать», чтобы уместить в оставшееся время. Мы обнаружили, что степень «сопротивления» любой активности такому сжатию может быть выражена одним параметром, который мы назвали упругостью (resilience) этой активности. В том случае, когда животное питается в течение обычного периода времени, но получает меньше пищи (рис. 24.27), упругость пищевого поведения относительно низка, поскольку оно не «сжимает» другие виды активности, несмотря на усилившийся голод животного. В том случае, когда животное добивается получения обычного количества пищи (рис. 24.28), упругость пищевого поведения относительно высока, поскольку ему удается потеснить другие виды активности в пределах общего времени, которым располагает животное; при этом само пищевое поведение ни в чем не страдает. Упругость поведения - этомера того, до какой степени может быть сжат во времени любой вид активности животного под влиянием других ее видов. Когда время является ограничителем бюджета, у животного наблюдается тенденция игнорировать те виды поведения, которые имеют низкую упругость. Действительно, если при ограничении времени какая-либо активность полностью исчезает из поведенческого репертуара животного, то мы можем назвать ее роскошью или праздной активностью. Непосредственные измерения упругости поведения почти не проводились, поскольку осуществить их практически очень трудно - для этого нужно в течение нескольких дней хронометрировать поведение животного в ситуации, когда экспериментатор может изменять количество времени, имеющегося в распоряжении животного. Такого рода исследование провел Крофт (Croft, 1975). Он изучил, как влияет длина светового дня на поведение и гормональ-
ный баланс самок канареечного вьюрка (Serinus canaria). Изменяя длину светового дня, Крофт тем самым изменял время, которым располагали канареечные вьюрки для осуществления разных видов активности, поскольку в темноте они не активны. Крофт обнаружил, что птицы тратят на питание одно и то же время как в короткие, так и в длинные дни, но в длинные дни они питаются более эффективно. В течение длинного дня птицы тратят больше энергии на различные другие виды активности, так что этот результат не столь уж неожидан. Птицы, содержащиеся на режиме длинного дня, тратят больше времени на сон и больше времени не заняты активной деятельностью, чем птицы, для которых установлен короткий день. Поскольку птицы имеют возможность спать ночью, представляется наиболее вероятным, что в условиях длинного дня птицы заполняют сном свободное время. Эту точку зрения подтверждает тот факт, что во время строительства гнезда птица меньше времени отводит на сон в дневное время. Крофт рассчитал, сколько времени остается у птицы на строительство гнезда за вычетом времени, необходимого для питания и чистки перьев (в расчет принималось и время перелетов с одного места на дру-
roe). Как видно из табл. 24.3, в длинные дни у птиц имеется достаточно времени для постройки гнезда, тогда как в короткие дни его очень мало. Однако интерпретация этих наблюдений осложняется тем обстоятельством, что длинные световые дни стимулируют выработку соответствующих гормонов, и поэтому в этих условиях мотивация строительства у птиц выше, чем в условиях короткого дня. Гнездостроителъное поведение птиц в условиях длинного дня более эффективно, чем в условиях короткого дня, и они тратят меньше времени на излишние повторения собирающих и собственно гнездостроительных движений. Таким образом, хотя понятие упругости поведения с теоретической точки зрения не связано с понятием мотивации (McFarland, Houston, 1981), при изучении временного бюджета животного трудно отделить влияние упругости поведения от влияния мотивации на это поведение. Однако упругость можно измерить косвенным путем и посредством функции спроса, которая используется экономистами для выражения связи между ценой тех или иных благ и их потреблением. Например, когда повышается цена на кофе, люди продолжают покупать его примерно столько же, сколько и раньше, - разве что чуть-чуть меньше (рис. 24.29). Однако стоит подскочить цене на фрукты, как спрос на них падает. Когда же повышается цена на свежую рыбу, спрос на нее снижается очень сильно. По всей вероятности, люди готовы платить большую цену, лишь бы не нарушить своей привычки пить кофе. Спрос на кофе называют неэластичным (inelastic). Однако, когда повышается цена на свежую рыбу, люди стараются покупать ее меньше или переключаются на другие продукты, например мясо или рыбные консервы. Спрос на рыбу называют эластичным. В поведении животных наблюдается явление, в точности аналогичное описанному. Если животное тратит определенное количество энергии на какую-то деятель- Таблица 24.3. Расчетные величины среднего времени, которым располагают для строительства гнезда овариэктомированные, но получающие эстроген птицы в условиях короткого (8 ч) и длинного (14 ч) светового дня, по сравнению со временем, которое необходимо для постройки гнезда
ность, то, как правило, оно будет меньше заниматься этой деятельностью, если энергетические траты на нее возрастут (рис. 24.30). Результаты многочисленных исследований показали, что функция спроса у животных подчиняется такому же общему закону, что и функция спроса у человека (Kagel et al., 1980). Например, Ли и Поупер (Lea, Poper, 1977) обнаружили, что спрос на пищу был эластичным у крыс, которым приходилось совершать определенную работу (нажимать на рычаг) для получения пищевого подкрепления. Если от крыс требовалось совершить больше работы (они должны были большее число раз нажимать на рычаг, чтобы получить одно подкрепление), то крысы продолжали работать, получая примерно такое же количество пищи. Однако эти исследователи обнаружили также и то, что эластичность спроса на пищевые шарики увеличивалась, если они были сделаны из сахарозы. Параллели между явлениями спроса у животных и человека - это тема, которая вызывает сейчас большой интерес (см., например, Allison, 1979, 1983; Lea, 1978; Rachlin, 1980). Эластичность функций спроса дает представление об относительной важности тех или иных благ (или деятельностей), на которые человек (или животное) тратит свои деньги (или энергию). Существует тесная взаимосвязь между эластичностью спроса и упругостью поведения (Houston, McFarland, 1980). Таким образом, функции спроса можно использовать как непрямую меру упругости. Если активность А имеет более высокую упругость, чем активность В, то функция спроса для активности А будет характеризоваться относительной неэластичностью, а для активности В - эластичностью (McFarland, Houston, 1981). ДЛЯ ЗАПОМИНАНИЯ 1. Стратегии пищедобывательного поведения - это сформировавшиеся в процессе эволюции стратегии, предназначенные для оптимизации определенных видов пищедобывательного поведения, с точки зрения связанных с ними затрат и выгод. 2. Многие виды поведения животных сбалансированы с другими поведенческими реакциями. Благодаря этому достигается компромисс между затратами и выгодами альтернативных вариантов активности. 3. Сумму всех возможных затрат и выигрышей можно выразить в виде функции затрат. Ее можно определить как мгновенный уровень риска, которому подвергается животное (и репродуктивных преимуществ, которые оно приобретает), когда оно находится в определенном внутреннем состоянии и занято определенной деятельностью в определенных условиях среды. 4. Можно допустить, что животные способны принимать рациональные решения в том смысле, что проявляемые ими предпочтения являются внутренне непротиворечивыми и обладают свойством транзитивности. 5. Существует определенная аналогия между экономикой потребления у человека и взаимоотношениями затрат и выигрышей, относящихся к поведению животных. Деньги аналогичны энергии, полезность аналогична выигрышу (увеличению приспособленности). На этой основе можно проанализировать бюджеты времени и энергии у животного. Рекомендуемая литература McFarland D.J., Houston A., 1981. Quantitative Ethology: The State Space Approach, London, Pitman. Heinrich A, 1979. Bumblebee Economics, Cambridge, Massachusetts, Harvard University Press.
|
|||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 502; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.011 с.) |