Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Функциональные аспекты смещенной активностиСодержание книги
Поиск на нашем сайте
Первоначально смещенную активность рассматривали как функционально неадекватную и аллохтонную (вызванную специфической энергией, не относящейся к делу). Обычно кажется, что поведение животного приводит к достижению цели, которая имеет очевидный биологический смысл. Поведение, которое не имеет очевидной цели, может показаться функционально неадекватным. Например, мы можем сказать, что целью пищевого поведения является поглощение пищи. Однако для смещенной пищевой активности у птиц характерно то обстоятельство, что они берут пищу в клюв, и затем роняют ее, а не проглатывают. Таким образом, смещенная пищевая активность будет казаться функционально неуместной, поскольку это пищевое поведение не выполняет своей нормальной функции. Однако может быть и так, что смещенная активность выполняет некоторые другие функции, например уменьшает мотивационное напряжение или облегчает коммуникацию с другими животными. Смещенная активность часто наблюдается при половых или агрессивных столкновениях, где, вероятнее всего, возникают мотивационные конфликты. Поскольку смещенная активность обычно имеет место в точке равновесия конфликтной ситуации, она, возможно, несет определенную информацию для другого участника столкновения. Например, мы уже видели, что у самца трехиглой колюшки начинают проявляться приступы смещенных копательных движений, когда он сталкивается с соперником вблизи границы своей территории, где тенденции нападения и избегания выравнены. Вполне возможно, что колюшки могли бы научиться узнавать территориальные границы по смещенному копанию, которое производят их соседи. В ситуациях, где такой тип коммуникации оказывается полезным, естественный отбор стремится сделать это поведение более эффективным и надежным в качестве источника информации. Эволюционный процесс, благодаря которому это происходит, носит название ритуализации. Ритуализация - это эволюционный процесс, благодаря которому определенные комплексы поведения модифицируются таким образом, чтобы осуществлять коммуникативную функцию. Эта концепция была намечена еще Джулианом Хаксли (Julian Huxley, 1923), однако развитие ее до полнокровной теории связано с именем Нико Тинбергена (Tinbergen, 1952). Данные, свидетельствующие о том, что ритуализация действительно возникла в ходе эволюции, были получены в сравни-
Рис. 21.8. Ритуализованная смещенная реакция чистки перьев. А. Пеганка (Tadorna tadorпа). Б. Кряква (Anas platyrhynchos). В. Чирок-трескунок (Anas querquedula). Г. Мандаринка (Aix galericulata). (Из The Oxford Companion to Animal Behaviour. 198Г.) тельных исследованиях. Например, считается, что чистка клюва, которую выполняют самцы зебровых амадин (Taeniopygia guttata) во время ухаживания, представляют собой смещенные действия. Обычно птица чистит клюв после еды, но при ухаживании чистка клюва наблюдается в точке равновесия между противоборствующими тенденциями приближения и избегания. У близких видов Lonchura punctulata и Lonchura striata никакой чистки клюва во время ухаживания не наблюдается - у них самец совершает поклон и в таком положении с опущенной головой остается на несколько секунд. Сходство этого движения с движением при чистке клюва и ситуация, в которой оно встречается, дают основание предположить, что этот поклон является ритуализованной формой чистки клюва. В процессе ритуализации формы поведения претерпевают различные изменения. Зачастую они становятся стереотипными и неполными. Ритуализованное поведение чистки тела, например, чаще всего ограничивается чисткой определенных частей тела. Движения становятся незавершенными и порой превращаются просто в символические акты. В других случаях движения становятся как бы замороженными, превращаются в определенные позы. Например, угрожающая поза самца трехиглой колюшки очень похожа на элемент его поведенческой реакции, когда он роет носом песок (см. рис. 21.3), и, вероятно, представляет собой ритуализованную форму смещенной активности рытья песка (Tinbergen, 1951). Действия, интенсивность которых в норме бывает весьма вариабельной, приобретают фиксированную интенсивность (Morris, 1957), как только становятся ритуализованными. Например, когда черный дятел долбит дупло для гнезда, ритмика его действий весьма нерегулярна. Самцы также барабанят по сухому дереву, чтобы привлечь самок и отпугнуть других самцов. В этом случае барабанная дробь отличается высокой стереотипностью и четким ритмом. Во время выдалбливания дупла для гнезда дятел прилетает к входному отверстию дупла и начинает в очень медленном ритме долбить по его краю. Эта медленная дробь - сигнал для другой птицы, приглашающий ее прилететь и продолжить работу по выдалбливанию дупла. Здесь мы имеем дело с нормальной активностью (выдалбливание гнездовой камеры) и двумя формами ритуализованных движений (когда дятел барабанной дробью оповещает о том, что данная территория занята, или призывает к смене «рабочих»). Эти ритуализованные сигналы очень четко отличаются один от другого и от нормального поведения. Для ритуализованного поведения зачастую необходимо развитие особых структур, или меток, которые усиливают демонстрационную картину, делая ее более выразительной и заметной. Лоренц (Lo- renz, 1941) отмечал, что чистка перьев у разных видов уток - это общая черта их ритуала ухаживания. У обыкновенной пеганки (Tadorna tadorna) это, по-видимому, неритуализованная смещенная активность. Она очень напоминает нормальную чистку перьев и проявляется во время конфликта мотиваций. Кряква (Anas platyrhynchos) поднимает одно крыло и производит чистку перьев лишь в том месте, где они образуют яркоокрашенную отметину; таким образом, птица как бы выставляет на обозрение эти перья (рис. 21.8). Это поведение выглядит ритуализованным в двух отношениях: 1) оно ограничено определенным участком тела и 2) его выраженность усилена за счет особых выделяющихся меток, обнажаемых во время соответствующей демонстрационной реакции. У чирка-трескунка (Anas querquedula) движения, связанные с чисткой перьев, еще менее завершенные и направлены на светло-голубые перышки, находящиеся на внешней стороне крыла. Такие движения не выполняют никакой функции, а просто имитируют ее. У мандаринок (Aix galericulata) чистка перьев во время ухаживания сильно ритуализована: клюв просто касается увеличенного ржаво-красного пера, которое поднимается вверх (рис. 21.8). Демонстрационные движения «чистки» как бы подчеркиваются хохолком, торчащим вдоль задней поверхности головы. При сравнении этих видов уток становятся заметными прогрессивная ритуализапия оригинальных смещенных действий и пути превращения этого поведения в эффективный и выразительный сигналы. ДЛЯ ЗАПОМИНАНИЯ 1. Смещенная активность требует специального объяснения, поскольку создается впечатление, что она проявляется вне связи со своей нормальной мотивацией. 2. Каким-то образом животные отвлекаются от выполняемого поведения и совершают серии кажущихся неадекватными действий. 3. Для того чтобы это произошло, поведенческая реакция высшего приоритета должна «выключиться» и тем самым растормозить смещенную активность. 4. Существует предположение, что некоторые типы смещенной активности в процессе эволюции становятся ритуализованными и, таким образом, начинают осуществлять коммуникативную функцию. Рекомендуемая литература Tinbergen N.. 1951, 1965. The Study of Instinct, Oxford, Clarendon Press.
Ритуализация и коммуникация Ранние этологи добились больших успехов в понимании коммуникативных связей у животных. Чарлз Дарвин (Darwin, 1872) сделал важный шаг на этом пути, когда указал на роль, которую играют в коммуникации внешние выражения эмоций у животных. Однако, как ни странно, в течение многих лет этим вопросом никто не занимался. Конрад Лоренц (Lorenz, 1932, 1935) использовал термин релизер для обозначения «тех характерных реакций, проявляемых представителями данного вида животных, которые активируют существующие пусковые механизмы у особей этого вида и вызывают у них определенные последовательности комплексов инстинктивного поведения». Таким образом, Лоренц фактически заложил основы классической этологической теории коммуникации, которая получила дальнейшее развитие в трудах Нико Тинбергена (Tinbergen, 1951, 1953). Различные специфические черты морфологии животного могут использоваться в ритуализованном поведении и действовать как сигнальные раздражители, в ответ на которые другие особи данного вида будут реагировать соответствующим образом. В случае социальных отношений эти сигнальные раздражители называют социальными релизерами. Например, Тинберген (Tinbergen, 1951) описывает, каким образом птенец серебристой чайки получает пищу от родителей. Когда родители, добыв корм, прилетают к гнезду, они начинают вызывать птенцов из укрытия. Птенец приближается к родителю и начинает клевать красное пятно на его клюве. Это заставляет родителя отрыгнуть пищу, которую он затем собирает с земли и держит в кончике клюва (рис. 22.1). Тинберген и др. (например, Tinbergen, 1949; Tinbergen, Perdeck, 1950) тщательно проанализировали характер стимулов, которые вызывают у серебристых чаек реакцию «выпрашивания» пищи. Используя ряд картонных моделей головы чайки, исследователи измеряли реактивность птенцов, которую выражали числом клеваний модели в течение определенного времени. Форму, окраску и расположение пятен на модели систематически изменяли (рис. 22.2). Эксперименты показали, что наибольшую реакцию вызывало предъявление моделей, которые располагались достаточно близко к земле, слегка двигались и обладали длинным, заостренным книзу выступом. Обычно птенцы нацеливаются на кончик клюва; реакция существенно усиливается, если на нем есть красное пятно, контрастно выделяющееся на общем фоне. Оказалось, что цвет клюва и головы не оказывает никакого влияния на реакцию выпрашивания пищи. Красное пятно на клюве серебристой чайки обладает всеми свойствами сигнального раздражителя. С точки зрения классической этологии оно «спускает реакцию выпрашивания пищи» у птенца. Этот тип социального релизера авторы рассматривали как основу социальной коммуникации у птиц: «Насколько позволяют судить наши современные знания, социальная кооперация, по-видимому, зависит главным образом от системы релизеров. Стремление животного подавать эти сигналы имеет врожденную основу, равно как и реактивность воспринимающего их. Релизеры, вероятно, всегда очень выразительны и относительно просты. Это очень важно, поскольку из других работ мы знаем, что стимулы, запускающие врожденное поведение, - это всегда простые сигнальные раздражители.
Рис. 22.1. Птенец серебристой чайки клюет красное пятно на клюве родителя. (Фотография Jim Shaffery.) Создается впечатление, таким образом, будто структуры и элементы поведения, выполняющие функции релизеров, приспособлены для того, чтобы служить сигнальными раздражителями» (Tinbergen, 1953). С точки зрения классических представлений считалось, что социальные релизеры являются специфическими для каждого биологического вида и развиваются как результат процесса ритуализации. Точно так же и их распознавание посредством врожденного пускового механизма считалось видовой характеристикой. Существовало представление, что коммуникационная система должна была развиваться таким образом, чтобы механизмы, которые посылают и воспринимают данный сигнал, были постоянно настроены друг на друга. В этой главе мы увидим, каким образом последующие исследования изменили это основное представление. Ритуализация В гл. 21 говорилось о том, что в процессе эволюции может происходить ритуализация смещенных действий и они, таким образом, начинают выполнять коммуникативную функцию. Однако не только смещенные действия, но и любые действия, которые являются потенциальным источником информации для других животных, могут превратиться в ритуализованные. Дарвин (1872) отмечал, что защитные мимические реакции у млекопитающих играют определенную роль в их коммуникации. Оборонительные рефлексы, в том числе сужение глаз, прижатие ушей и вздыбливание волос на шее, призваны защитить органы чувств в момент опасности. Такие реакции служат источником информации для других животных, которые могут интерпретировать ее как сигналы страха или гнева. Таким образом, примитивные мимические реакции дают хороший материал для отбора действенной коммуникативной системы. Эффективность этих выразительных реакций может быть повышена за счет их преувеличения, сопровождения специфической вокализацией и при помощи характерных отличительных «меток», привлекающих внимание к морде или подчеркивающих изменения мимики животных. Отсутствие волос на значительной части лица человека привлекает внимание к главным его чертам, которые используются при коммуникации (рис. 22.3). Дарвин заметил, что противоположные по значению сигналы зачастую передаются посредством выразительных реакций или поз, которые также противоположны по своему характеру. Мимические реакции человека, испытывающего удовольствие или гнев, осуществляются с помощью антагонистических комплексов мышц, а поза рассерженной собаки во многих отношениях противоположна позе дружелюбно настроенного животного (рис.
Рис. 22.2. Модели, которые использовались при экспериментальном исследовании реакции выпрашивания пищи у птенцов серебристой чайки. Длина цветной полоски пропорциональна числу клевков в ответ на предъявление данной модели. А. Модели, отличающиеся по окраске клюва. Б. Модели, отличающиеся по цвету пятна на желтом клюве. В. Модели, у которых клюв и пятно на нем были серыми, но различались по контрасту. (По Tinbergen, Perdeck, 1950.) 22.4). Дарвин (1872) назвал это принципом антитезы. Считается, что оборонительные реакции и их антитезы имели особенно важное значение в эволюции выразительной мимики у приматов (Andrew, 1963). Другой аспект поведения, который, по всей вероятности, обеспечил важное начало в развитии ритуализованных демонстраций, - это движение намерения. Оно представляет собой незавершенный поведенческий комплекс, который несет потенциальную информацию о том, что животное собирается совершить определен-
Рис. 22.3. Иллюстрации из книги Ч. Дарвина «Выражение эмоций у человека и животных».
Рис. 22.4. Дружелюбная (слева) и агрессивная (справа) позы собаки как иллюстрация дарвиновского «принципа антитезы». ные действия. Так, например, когда птица собирается взлететь, она сначала приседает, поднимает хвост и вытягивает голову (рис. 22.5). Прежде чем взлететь, птица может сделать несколько приседаний или не приседать вовсе. Сколь важную роль играют движения намерения в качестве сигналов для других животных, можно понять из работы Девиса (Davis, 1975) по изучению поведения полета у голубей. Он обнаружил, что голубь, покидающий стаю, не вызывает беспокойства у других птиц при условии, что он проделывает обычные движения, демонстрирующие намерение взлететь. Если же голубь взлетает внезапно, без каких-либо предварительных сигналов намерения, тогда и все птицы поднимаются в воздух. По-видимому, взлет, которому не предшествовали движения намерения, является своего рода сигналом тревоги. Изучением ритуализованных движений намерения занимались многие этологи (например, Daanje, 1950; Tinbergen, 1953; Andrew, 1956a). Такие движения, например, проявляются во время ухаживания у американской зеленой кваквы и желтоглазой утки. Американская зеленая кваква, которую изучал Мейеррикс (Meyerriecks, 1960), строит гнезда на мертвых деревьях в соленых маршах. Самцы, прилетевшие весной с мест зимовки, начинают охранять гнездовые деревья от самцов-соперников. Пришельца встречают демонстративным поведением угрозы (рис. 22.6,А), которое, скорее всего, возникло как результат эволюции поведения, связанного с намерением нападения. При этом самец принимает горизонтальную позу, направляет клюв в сторону соперника, распушает перья и начинает вибрировать хвостом. Самок привлекают призывные сигналы самцов, но как только самки приблизятся к самцам, последние начинают угрожать и самкам. По мере проявления самками настойчивости поведение самца изменяется. О своей готовности принять самку он будет сигнализировать ей посредством особой позы, сопровождаемой специфическим щелканьем (рис. 22.6, Б). Эта поза выражается в том, что клюв птицы по диагонали направляется вниз, и производит характерные щелкающие звуки. Такая демонстративная поза напоминает поведение, при котором самец отламывает от дерева веточки, необходимые ему для постройки гнезда. Не исключено, что эта демонстрация является ритуализованной формой смещенного гнездостроительного поведения. После того как самец примет самку, он принимает демонстративную позу «вытягивания шеи», которая, по-видимому, представляет собой ритуализованную
Рис. 22.5. Многие птицы приседают и приподнимают хвост перед тем, как взлететь.
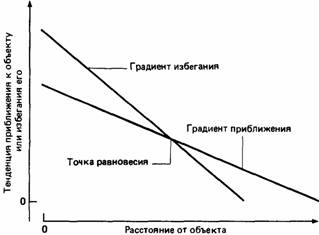
форму намерения взлететь. Во многих отношениях эту позу можно рассматривать как своеобразную антитезу другого поведения, выражающегося в устремленной вперед позе угрозы. Если эта последняя сопровождается резкими звуками и распушением оперения, которое как бы увеличивает размеры птицы, то поза вытягивания шеи, напротив, сопровождается мягкими звуками и прижатием («прилизыванием») перьев. Во время демонстрации угрозы клюв - главное оружие птицы - направляется на соперника, в результате чего обнажается ярко-красное очертание рта. Когда самец вытягивает шею, он направляет клюв в сторону от самки. Если устремленная вперед поза означает угрозу, то поза вытягивания шеи символизирует умиротворение. За этой позой следуют взаимные демонстрации со стороны самки и самца, которые проявляются в виде «целования» клювами и взаимной чистки оперения. Очень скоро после установления такого контакта происходит копуляция. Конфликт Как правило, отправной точкой для процесса ритуализации служит конфликт мотиваций. Он наблюдается в том случае, когда в регуляции поведения начинают конкурировать за доминирование две тенденции (см. гл. 25). Поскольку конфликтующие тенденции не могут проявиться в поведении одновременно, поведение во время конфликта очень отличается от обычного гладкого проявления поведенческой активности. Исходя из логических соображений, можно представить себе три основных типа конфликта, хотя практическое значение из них имеет только один. 1. Конфликт приближение-приближение возникает тогда, когда две одновременные тенденции направлены на различные цели. Хотя у животного и может наступить такое состояние, когда эти тенденции будут выравнены, оно бывает очень непродолжительным, поскольку любое отклонение от точки равновесия приведет к усилению одной из тенденций приблизиться к своей цели. Такая нестабильность объясняется существованием градиента цели (рис. 22.7), благодаря которому тенденция приблизиться к цели возрастает по мере сокращения расстояния до нее. 2. Конфликт избегание-избегание возникает в том случае, когда одновременно наблюдаются две тенденции избегания. Поскольку тенденция избежать каких-либо объектов обычно усиливается пропорционально близости к объекту (см. рис. 22.7), животное стремится занять такое положение, где эти тенденции избегания выравниваются, а затем оно старается выйти из этой ситуации, удаляясь под прямым углом к линии, соединяющей эти два объекта. Поэтому ситуация избегание-избегание является нестабильной. 3. Конфликт приближение-избегание возникает тогда, ко-
Рис. 22.6. Демонстрационное поведение у зеленой кваквы (Butorides virescens). А. Агрессивная демонстрация устремления вперед. Б. Демонстрация «щелканье клювом». В. Демонстративная поза с вытянутой шеей. (Из The Oxford Companion to Animal Behaviour, 1981.) Рис. 22.7. Целевые градиенты для реакций приближения и избегания.
гда у животного одновременно появляются тенденции приблизиться к объекту и убежать от него. Как видно из рис. 22.7, в такой ситуации градиенты приближения и избегания обычно пересекаются. Это означает, что около объекта тенденция избегания сильнее тенденции приближения, а когда животное находится вдали от объекта, наблюдается обратная ситуация. Где-то между этими позициями находится точка равновесия, в которой тенденции приближения и избегания одинаковы. Таким образом, конфликт приближение-избегание имеет четкую тенденцию к стабильности, поскольку животное будет двигаться по направлению к точке равновесия, где бы оно ни находилось изначально. Конфликт приближения и избегания - очень распространенное явление в поведении животных. Так, например, самец трехиглой колюшки пытается справиться с таким конфликтом (между страхом и агрессией по отношению к самцу-сопернику), когда находится вблизи границы своей территории. При ухаживании также возникает конфликт приближения и избегания, поскольку каждое животное сначала испытывает чувство настороженности по отношению к другому животному, хотя и привлекающему его в сексуальном отношении. Теоретически в точке равновесия конфликтной ситуации животное должно оказаться в тупике, поскольку, куда бы оно ни двигалось, тенденции приближения и удаления будут приводить его обратно к точке равновесия. Однако практически в конце концов побеждает какая-то одна тенденция, либо потому что будет уменьшаться страх животного перед новым объектом, либо потому что будет усиливаться его стремление приблизиться к этому объекту, либо потому что в дело вмешается какой-то другой мотивационный фактор. Тем не менее можно наблюдать, как во время конфликта приближение-избегание животное то приближается к объекту, то удаляется от него или подолгу остается в амбивалентной позе. Поведение, которое состоит из отдельных компонентов конфликтующих тенденций, называется компромиссным поведением. Конфликт может проявиться и в том, что животное принимает амбивалентную позу, в которой смешиваются элементы этих конфликтующих тенденций (Andrew, 1956с). Так," например, когда люди в парке предлагают утке хлеб, она может приблизиться, а затем остановиться, вытягивая шею вперед, чтобы схватить хлеб, и в то же время поворачивать тело назад. Как мы уже видели в гл. 21, для конфликтных ситуаций типична смещенная активность; считается, что такая активность возникает в точке равновесия конфликтующих тенденций. Мотивационное состояние животного в конфликтной ситуации обычно проявляется очень наглядно. Таким образом, это идеальный материал для ритуализации. Многие демонстрации, по-видимому, состоят из ритуализованных компонентов конфликтного поведения, и многие из них анализировались с этой точки зрения. Например, у серебристой чайки вертикальная поза угрозы (рис. 22.8) содержит элементы и страха, и агрессивности (Tinbergen, 1959). С одной стороны, прижатые перья, вытянутая вверх шея и поворот боком к противнику - это признаки страха. С другой, направленный вниз клюв и поднятые карпели крыльев - это уже признаки агрессивности. Интерпретировать демонстрационные позы, исходя из их эволюционной природы и значения, в настоящее время очень трудно. Тем не менее этологи добились поразительных успехов при анализе различных типов поведения, проявляемого при угрозе и ухаживании, с точки зрения противоборства относительно небольшого числа поведенческих тенденций. Обычно исходят из того, что существуют три основные тенденции, одна из которых, если действует изолированно, приводит к половому поведению, другая - к атаке, а третья - к убеганию. Эти виды активности очень редко проявляются в чистом виде. Наблюдаемое поведение интерпретируют как результат смешения этих трех основных тенденций. Так, например, поза угрозы обычно представляет собой определенную комбинацию тенденций атаки и бегства. Такой подход был использован для анализа различных видов активности, например ухаживания, церемонии смены родителей на гнезде и поведения группового отпугивания хищников, наблюдающегося у мелких птиц. Признание того факта, что в поведении проявляется конфликт, и анализ этого поведения с точки зрения лежащих в его основе несовместимых тенденций составляют сущность подхода, который получил известность как «конфликтная теория демонстраций» (Baerends, 1975). Наиболее отчетливо этот подход проявился в работах Тинбергена (Tinbergen, 1959; 1962) и его коллег. Рассматривая демонстрации животных с точки зрения конфликта тенденций, необходимо учитывать четыре категории данных (Tinbergen, 1962; Hinde, 1966): 1. Ситуация. Например, вблизи границы своей территории животное, скорее всего, будет и испытывать страх, и про- являть агрессивность, а в присутствии потенциального полового партнера в конфликте может участвовать и половая мотивация. 2. Поведение животного во время демонстрации. Это может быть явное конфликтное поведение, например попеременное приближение к сопернику и удаление от него. Но это могут быть и изменения окраски животного, коррелированные с его мотивацией. Так, например, изменения характера окраски самца гуппи указывают на выраженность его половой мотивации. 3. Поведение, которое непосредственно предшествует демонстрации или же следует за ней. Мойниган (Moynihan, 1955) использовал этот метод для оценки относительной силы тенденций атаки и убегания, которые проявлялись в различных демонстрациях у обыкновенной чайки (рис. 22.9). Однако при использовании этого метода возникает целый ряд трудностей. Во-первых, поведение, которое проявляется сразу после демонстрации, может оказаться реакцией на ответ другого животного на эту демонстрацию. Эту трудность можно обойти, если анализировать только то поведение, которое проявляется при неизменном поведении соперника. Можно также проверять теории, касающиеся демонстраций, используя неподвижные модели животных-соперников. Во-вторых, поведение, которое предшествует демонстрации и сразу следует за ней, может быть результатом действия целого комплекса мотивационных факторов, а не выражением одной
Рис. 22.8. Вертикальная поза угрозы у серебристой чайки. (По фотографии Тинбергена.)
Рис. 22.9. Позы угрозы у обыкновенной чайки. Различные позы связаны с различным соотношением тенденций к атаке (темные столбцы) и избеганию (светлые столбцы). (По Moynihan, 1955.) простой тенденции. И наконец, в-третьих, поведение может быть мотивациоино ие связано с данной демонстрацией, как это бывает в случаях смещенной активности и при организации поведения по принципу разделения времени (см. разд. 25.4). 4. Природа демонстрации. Каждую позу можно проанализировать с точки зрения ее компонентов (угол наклона головы, конечностей и т. д.). Например, Тинберген (Tinbergen, 1959), исследуя поведение серебристой чайки, проводит различие между позой агрессивной угрозы (см. рис. 22.8) и позой угрозы у испуганной птицы; в этом последнем случае клюв располагается более горизонтально. Эдвина Баер и я показали, что положение перьев на различных участках тела агрессивно настроенных или защищающихся горлиц были типичны именно для таких состояний и отличались от положения перьев, вызванных изменениями температуры (рис. 22.10) (McFarland, Edwina Baher, 1968). Для подтверждения данных такого рода иногда может быть полезен сравнительный анализ поведения животных различных видов. Например, во время драки серебристые чайки бьют крыльями своих соперников; элементом их угрожающей вертикальной позы является приподнимание карпелей крыльев, как будто птицы готовятся к драке. Поморники, напротив, во время драк не ударяют соперников крыльями, и поэтому в их вертикальной позе угрозы нет демонстрации карпелей. Данные этого типа зачастую можно использовать для того, чтобы дать достоверное объяснение мотивации, лежащей в основе демонстраций. Однако иногда в этой процедуре есть элемент «движения по кругу». Обстоятельные данные, полученные в полевых исследованиях, в ряде случаев нашли экспериментальное подтверждение. Хайнд (Hinde, 1952) описал различные позы угрозы, которые он наблюдал во время территориального конфликта на границе территорий у большой синицы (Parus major). Все эти позы представлены на рис. 22.11. Хайнд сформулировал гипотезу о том, что демонстрации угрозы возникают при одновременном действии побуждений к атаке и бегству. Блёртон-Джонс (Blurton-Jones, 1968) попытался проверить эту гипотезу в экспериментах, где можно было независимо друг от друга регулировать тенденции атаки и бегства. Сначала Блёртон-Джонс наблюдал за птицами, находящимися в неконтролируемых ситуациях, и использовал методы анализа, которые были описаны выше. В результате весьма тщательного анализа он пришел к выводу, что демонстрации угрозы возникают по тем же причинам, что и атака, и что факторы, обычно вы-
Рис. 22.10. Положение перьев у горлицы (Streptopelia risoria). Чтобы получить показатель положения перьев, тело птицы разделяли на ряд областей и один раз в минуту отмечали положение перьев в каждой области. Разным положениям перьев приписывали разные оценки, и эти оценки суммировали по десяти тестам, так что в итоге могло получиться от 0 (полностью приглаженное оперение) до 20 баллов (полностью взъерошенное оперение). Исследование проводили, когда птица сидела на гнезде. Во время эксперимента деревянную палочку каждые 2 мин пододвигали примерно на 50 мм ближе к птице. Горлица реагировала на это оборонительной позой, для которой были характерны вполне определенные изменения положения перьев на различных областях тела: 1 -верхняя часть головы; 2-шея; 5-область спины; 4-область живота; 5-крыло; 6-грудь. (По McFarland, Baher, 1968.)
Рис. 22.11. Позы большой синицы (Parus major), которые можно наблюдать во время территориальных конфликтов. А - обычная расслабленная поза, которая постепенно переходит в демонстрацию угрозы с опущенной головой (Б-Г) или в демонстрацию угрозы с поднятой головой (Д-Ж). Горизонтальная демонстрация (3, И) иногда сопровождается демонстрацией с раскрыванием крыльев (К, Л). (По Blurton-Jones, 1968.) зывающие бегство, оказывают мало влияния на характер этой демонстрации. Он постулировал, чтодемонстрации угрозы наблюдаются у животных тогда, когда налицо преобладание факторов, вызывающих атаку, но осуществлению атаки препятствуют тенденция к бегству или какие-либо другие факторы. В том случае, когда у птицы наблюдалась тенденция оставаться на одном месте, связанная со стремлением получать там пищу, это проявлялось в увеличении количества «горизонтальных» демонстраций в отличие от демонстраций с поднятой или опущенной головой. Таким образом, создается впечатление, что характер демонстрации угрозы определяется обстоятельствами, препятствующими осуществлению атаки. При проведении экспериментов избирательно (независимо друг от друга) изменяли величину тенденций к атаке, бегству и питанию. Атака, которую вызывали у птицы легкими ударами кончиком карандаша, сопровождалась следующими позами: позой с опущенной головой, позой с горизонтально вытянутой головой, позой с поднятой головой, раскрыванием крыльев. Бегство птицы можно было вызвать с помощью маленькой лампочки; при этом птица поднимала «хохолок» и распушала перья. Когда птице предлагали пищу, она делала прыжок вперед, тянулась к пище и схватывала ее. Предъявляя эти стимулы одновременно. Блёртон-Джонс имел возможность искусственно создавать конфликтные ситуации. Одновременное предъявление стимулов для атаки и бегства создавало конфликт атака-бегство, в котором движения атаки были существенно редуцированы, а поза угрозы с поднятой головой и другие угрожающие позы были заметно более выражены. Таким образом длительные атаки можно было трансформировать в демонстрации угрозы, предъявляя птице стимулы бегства с внешней стороны клетки, находясь в которой птица не могла спасаться бегством. Эти данные подтверждают выводы, сделанные на основе наблюдений за поведением птиц в природе: любой сигнал, который останавливает атаку при наличии стимула атаки, будет вызывать у животного демонстрацию угрозы. Блёртон-Джонс пришел к заключению, что такого результата нельзя было ожидать, если бы демонстрации угрозы представляли собой неритуализованные комбинации компонентов атаки и бегства. Вместе с тем демонстрации могут быть ритуализованы до такой степени, что на них уже не будет сказываться влияние тенденции к бегству; они будут связаны только с тенденцией к атаке. Вполне возможно, что какая-то демонстрация может полностью потерять связь с обусловившими ее изначально факторами и у нее появится свой собственный причинный фактор. Однако в демонстрациях угрозы большой синицы этого, по-видимому, не наблюдается. Как результаты наблюдений, так и экспериментальные данные показывают, что демонстрации угрозы имеют по существу ту же мотивацию, что и поведение атаки, хотя форма проявления демонстрации зависит от конкретных обстоятельств.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 716; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.129.194.30 (0.013 с.) |