Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
II. 1. Восприятие у животныхСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте В этих трех главах мы рассматриваем нервную и сенсорные системы животных. В главе 11 описаны основные элементы нервной, сенсорной и мышечной систем. Затем следует обзор нервных систем безпозвоночных и позвоночных животных. Мы стремимся дать общую картину физиологического аппарата, характерного для разных видов всего животного царства. В гл. 12 рассмотрены главные сенсорные системы, включая хеморецепцию, слух и зрение. Обсуждаются также три разных подхода к восприятию. Гл. 13 посвящена экологическим аспектам сенсорных систем, в особенности специальным сенсорным адаптациям животных, обитающих в неблагоприятных средах.
Иоганнес Мюллер (1801-1858)
Иоганнес Петеp Мюллер, сын бедного сапожника, положил начало экспериментальной физиологии в Германии и в значительной степени определил наше представление об организме как о машине. У Мюллера было много знаменитых учеников, в том числе Дюбуа-Реймон. Гельмгольц, Генле, Кёлликер, Ремак. Рейхерт и Вирхов. Он был в числе пионеров многих разделов биологии, в частности сенсорной физиологии и морской зоологии. Им опубликована монументальная книга «Handbuch der Physiologie des Menschen fur Vorlesungen» («Руководство для лекций по физиологии человека»), переведенная на английский язык У. Бейли и изданная в 1827 г. под названием «Elements of Physiology» («Элементы физиологии»). Самым важным вкладом Мюллера в изучение поведения является его доктрина «специфических нервных энергий». До этого считали, что каждое отдельное событие в окружающей среде (стимул) действует на сенсорные нервы специфическим для этого стимула образом. Мюллер открыл, что данный нерв всегда производит один и тот же тип ощущения независимо от способа стимуляции. Так, падающий на глаз свет вызывает зрительное ощущение, но такой же эффект производит и механическое раздра- жение, например удар по глазу, а также электрическое раздражение зрительного нерва. Если бы можно было соединить со зрительным нервом наше ухо, то звуковая стимуляция тоже вызывала бы зрительное ощущение. Таким образом, важен не орган чувств, а нервы, передающие сенсорные сообщения тем частям головного мозга, которые получают зрительную, слуховую, тактильную и обонятельную информацию и переводят ее в соответствующие ощущения. Органы чувств преобразуют различные формы внешней стимуляции - например, свет, тепло и механическую энергию в электрический потенциал, который могут зарегистрировать рецепторные клетки, т. е. клетки, составляющие часть органа чувств и связанные с другими клетками нервной системы. Доктрина специфических нервных энергий - основной организующий принцип сенсорной физиологии. Она устраняет все прежние спекуляции относительно роли органов чувств в восприятии, ясно показывая, что они прежде всего являются преобразователями одной формы энергии в другую. Хотя некоторая фильтрация стимула может происходить на периферии, именно головной мозг сортирует и классифицирует поступающую информацию, исходя только из интенсивности стимуляции периферических сенсорных нервов. Отчасти благодаря трудам Мюллера и его учеников Дюбуа-Реймона и Гельмгольца стало известно, что импульсы, бегущие по определенному аксону, обладают измеримой величиной и скоростью распространения, характерными именно для данной нервной клетки; это означает, что она может передавать сообщения только об интенсивности стимуляции. Другую информацию о природе стимула мозг может получать, только интегрируя информацию, поступающую от многих нервных клеток.
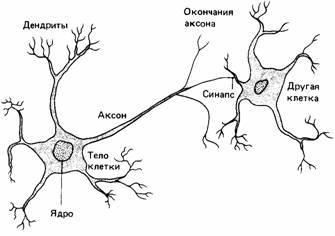
Нервная регуляция поведения В этой главе мы рассмотрим нервную регуляцию поведения, начиная с изложения ее общих принципов; затем дадим обзор типов нервной системы, встречающихся у животных, и проследим, как общая организация нервной системы соотносится с поведением. Нервные клетки Нервная система состоит из нервных клеток, называемых нейронами, которые специализированы для передачи информации от одной клетки другой. У каждого нейрона имеется тело с ядром и множество ветвящихся отростков (рис. 11.1). Обычно у клетки много коротких отростков, называемых дендритами, и один длинный - аксон. Дендриты образуют связи с близлежащими нейронами, а аксоны проводят сигналы на сравнительно большие расстояния. Мембрана нейрона обычно поляризована. Иными словами, между ее наружной и внутренней поверхностью имеется электрический потенциал, который в неактивном нейроне называется потенциалом покоя и создает устойчивое состояние готовности, сходное с состоянием электрической батареи, накопившей энергию, которая высвобождается в случае надобности. Потенциал покоя обусловлен неодинаковыми концентрациями ионов К + внутри и снаружи клетки. Когда клетка находится в состоянии покоя, внутренний заряд отрицателен по отношению к наружному. При деполяризации клетки ее мембранный потенциал снижается по направлению к нулю (рис. 11.2). Когда же мембранный потенциал становится более отрицательным, говорят, что клетка «гиперполяризована». Если потенциал покоя падает ниже определенного порогового значения, вдоль мембраны распространяется потенциал действия. Он длится недолго и вызывается закономерными изменениями относительных концентраций ионов Na+ и К+ по обе стороны мембраны (рис. 11.3). Потенциал действия проходит к концу аксона в виде электрической волны. Он всегда имеет одну и ту же амплитуду (высоту), обычно зависящую от диаметра аксона. По более крупным аксонам распространяются более высокие потенциалы действия (и с большей скоростью), чем по более тонким аксонам. После прохождения каждого потенциала возникает рефрактерный период, в течение которого мембрана восстанавливает свое нормальное ионное равновесие и нормальный потенциал покоя. Поскольку во время рефрактерного периода новый потенциал действия возникнуть не может, рефрактерные свойства аксона определяют максимальную частоту потенциалов действия. Когда возникает потенциал действия, говорят, что нейрон «разряжается». Этот потенциал часто проявляется в виде пика (спайка) на экране осциллографа, приспособленного для измерения мембранных потенциалов с помощью электродов, введенных в нервную ткань. Нейрон разряжается по закону «все или ничего» (или возникает полный спайк, или же его совсем нет), причем частота разряда ограничена рефрактерными свойствами нейрона
и зависит от силы его стимуляции. Таким образом, сообщение, которое посылает нейрон, кодируется частотой (рис. 11.4). Мембраны аксонов и дендритов не образуют физических связей с другими нейронами, а очень близко подходят к ним в соединениях. называемых синапсами. Обычно в синапсе выделяются очень малые количества химических нейромедиаторов, которые влияют на потенциал покоя воспринимающей мембраны и, следовательно, на готовность воспринимающего нейрона генерировать потенциалы действия. Нейроны могут быть стимулированы другими нейронами, повреждением или сенсорными рецепторами. Во всех случаях принцип один и тот же. Стимуляция вызывает изменение мембранного потенциала, и когда он достигает порогового уровня, генерируется потенциал действия. Теперь рассмотрим, как этот процесс происходит в сенсорных рецепторах. Сенсорные рецепторы Сенсорные рецепторы - это специализированные (часто нервные) клетки, ответственные за преобразование и передачу информации. Как и обычные нервные клетки, они имеют дендриты и один или более аксонов. Рецепторы специализированы в соответствии с той энергией среды. на которую они реагируют. Например, фоторецепторы содержат пигменты, которые химически изменяются под действием света, и при такой стимуляции возникает электрический потенциал. В механорецепторах происходят электрохимические изменения вследствие деформации мембраны клетки. Преобразование энергии обычно совершается в теле клетки, и для всех рецепторов характерно, что энергия окружающей среды превращается в градуальный электрический потенциал, называемый генераторным потенциалом, который обычно пропорционален интенсивности стимуляции рецептора. Когда генераторный потенциал достигает определенного порогового уровня, он запускает потенциал действия, который бежит по
аксону рецепторной клетки. В этом заключается передаточная часть сенсорного процесса, причем информация обычно кодируется так, что, чем сильнее стимул, тем выше частота потенциалов действия. В отсутствие стимуляции генераторный потенциал постепенно снижается до уровня покоя. Когда он падает ниже порогового значения, потенциалы действия перестают генерироваться. При возобновлении стимуляции может возникнуть короткая задержка (латентный период), пока генераторный потенциал возрастает от уровня покоя до порогового. При прерывистой стимуляции он ритмически повышается и понижается, генерируя залпы потенциалов действия. Однако, если частота прерывистой стимуляции достаточно высока, генераторный потенциал может не успевать снизиться в перерывах между стимулами, и тогда генерация потенциалов действия станет непрерывной. Этим объясняется то, что при очень высокой частоте прерывистой стимуляции мы не способны отличать ее от непрерывной. Этот феномен слияния мельканий присущ всем органам чувств, что наиболее очевидно в случае зрения. Тот факт, что быстро мелькающий свет вызывает такое же зрительное ощущение, что и постоянный, делает возможным телевидение и кино. Потенциалы действия, передающие сенсорную информацию, ничем не отличаются от любых других нервных импульсов. Их величина определяется размерами аксона, а частота - силой стимуляции. Каждый тип рецепторов посылает импульсы прямо или опосредованно в определенный отдел мозга. Испытываемые ощущения зависят не от типа рецептора или сообщений, которые он посылает, а от той части
мозга, которая принимает эти сообщения. От головного мозга зависит также локализация ощущения. Так, например, при боли нервные волокна от кисти посылают сигналы в одну часть мозга, от предплечья - в другую и т.п. «Боль», испытываемая мозгом, локализуется в той части тела, откуда пришло сообщение. Это явление иллюстрируется сообщениями людей, перенесших ампутацию конечностей. которые жалуются на болевые ощущения, идущие, как им кажется, от удаленной (фантомной) конечности. Раздражение перерезанных нервных окончаний посылает импульсы в те части мозга, которые были связаны с ампутированной конечностью. Мозг истолковывает поступающие сигналы как идущие от утраченной конечности, и возникающие ощущения зависят от того, какой нерв раздражен. От такой фантомной конечности могут приходить также ощущения тепла, холода или прикосновения. Мышцы и железы Нервная система управляет поведением и до некоторой степени внутренней средой животного (см. гл. 15). Это управление производится приказами, отдаваемыми мышцам и железам. В мышечных клетках имеются сложные белковые молекулы, способные к сокращению и расслаблению. Нервные окончания связаны с мышцами через синапсы, сходные с теми, какими соединены друг с другом нейроны. Придя в нервно-мышечное соединение, нервные импульсы вызывают электрические потенциалы, заставляющие мышцу сокращаться. Ее расслабление возникает при отсутствии стимуляции. Сокращаясь, мышца укорачивается, если этому не препятствует удерживание обоих ее концов. При расслаблении мышца может удлиниться, но только если ее растягивают другие мышцы или какая-нибудь внешняя сила. Мышцы обычно расположены антагонистическими, противодействующими друг другу группами. У некоторых беспозвоночных, например у кольчатых червей, мышечному сокращению может препятствовать гидростати-
ческое давление, повышающееся при сжатии мускулатурой части полости тела. Это давление заставляет мышцы удлиняться при расслаблении. У других беспозвоночных, например членистоногих, мышцы находятся внутри жесткого наружного скелета, который образует не- обходимую систему рычагов для антагонистических групп мышц (рис. 11.5). У позвоночных животных такой системой служит внутренний скелет, а мышцы расположены так, что тянут его части в противоположные стороны (рис. 11.6). Одна группа мышц расслабляется, когда другая сокращается. Некоторые железы находятся под нервным контролем. У позвоночных к ним относятся, например, слюнные железы, мозговая часть надпочечников, вырабатывающая адреналин, и задняя доля гипофиза, в которой образуется несколько важных гормонов. Секреты этих желез могут влиять на поведение косвенно, воздействуя на внутреннее состояние животного, как будет показано в конце этой главы. Соместетическая система Головному мозгу животного важно получать информацию о состоянии организма. За положением конечностей, давлением на внутренние органы, температурой разных частей тел и многими другими свойствами следит центральная нервная система (ЦНС) посредством внутренних рецепторов (интероцепторов), расположенных в «стратегически важных» пунктах. Эта система, ответственная за телесные ощущения, называется соместетической. В коже, скелетных мышцах1 и внутренних органах позвоночных находится множество типов рецепторов. Некоторые из них показаны на рис. 11.7. Беспозвоночные также имеют широкий диапазон рецепторов. Человек обладает пятью типами кожных рецепторов, вызывающих ощущения прикосновения, давления, тепла, холода и боли. Болевых рецепторов много, в 27 раз больше, чем Холодовых, и в 270 раз больше, чем тепловых. Некоторые кожные рецепторы отличаются быстрой сенсорной адаптацией. В ответ на ступенчатое изменение стимуляции частота нервных импульсов быстро повышается, а затем снижается до уровня покоя. Это значит, что рецептор служит хорошим индикатором изменений в силе стимуляции, но плохим индикатором ее абсолютного уровня. Это дает преимущества в тех случаях, когда от кожных рецепторов требуется быстрая информация об изменениях среды, которые могут подействовать на организм, например об изменениях температуры. Рецепторы, расположенные глубоко в теле, выполняют множество разнообразных функций, в том числе отмечают изменение кровяного давления, напряжение мышц, количество соли в крови и т. д. Мы не осознаем непосредственно информацию, передаваемую большинством интероцепторов. Они не вызывают ощущений. Иногда их действия комбинируются, давая ощущения голода, жажды или тошноты, но это происходит вследствие сложных процессов в мозгу, которые не всегда связывают ощущение с определенными частями тела. Вероятно, это объясняется тем, что действие, которое должно быть произведено в ответ на голод и жажду, является гораздо менее непосредственным, чем ответ на прикосновение или поверхностное изменение температуры. Ориентация животных по отношению к силе тяжести или внешним стимулам, подобным свету, отчасти зависит от информации о пространственном соотношении разных частей тела. У млекопитающих такая информация поступает от вестибулярной системы и рецепторов в суставах, мышцах и сухожилиях. Суставные рецепторы дают информацию об угловом положении каждого сустава (Howard, Templeton, 1966). В сухожилиях млекопитающих заложены сухожильные рецепторные органы Гольджи, чувствительные к напряжению. Они посылают сигналы в спинной мозг и участвуют в простом рефлексе, который противодействует повышению мышечного напряжения. В мышцах находятся мышечные веретена, чувствительные к изменениям длины мышцы (рис. 11.8). Они состоят из видоизмененных мышечных волокон, вокруг средней части которых закручено спиральное нервное окончание, называемое
1 Рецепторы в скелетных мышцах обычно объединяют в кинестетическую систему. - Прим. перев.
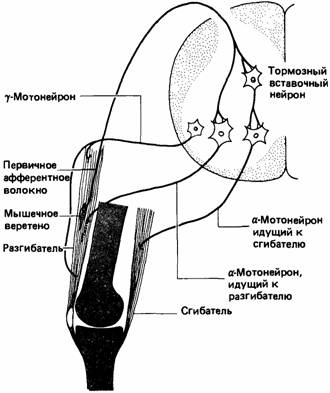
первичным (или аннулоспиралъным) окончанием. При удлинении мышцы веретено растягивается и посылает быстрые сигналы в спинной мозг. Встречаются также вторичные гроздевидные окончания, посылающие более медленные сигналы. Во многих веретенах млекопитающих имеются и первичные, и вторичные окончания, а в других - только первичные (Prosser, 1973). Эти веретена участвуют в простом рефлексе, который противодействует удлинению мышцы. Мышечные веретена заключены в фузиформную соединительную ткань, и их мышечные волокна называются интрафузальными в противоположность обычным экстрафузальным. Последние иннервируются альфа-мотонейронами, тела которых находятся в спинном мозгу. У млекопитающих интрафузальные волокна интернируются более мелкими гамма-мотонейронами, поддерживающими веретено в тоническом состоянии активности, благодаря чему для его активации требуется меньшее мышечное растяжение. Поскольку мышечные веретена расположены параллельно экстрафузальным волокнам, при сокращении мышцы они склонны к расслаблению. Гамма-нейроны могут приказать интрафузальным волокнам напрячься, в результате чего веретено сохранит состояние готовности (рис. 11.9). У птиц мышечные веретена напоминают веретена млекопитающих и экстра- и интрафузальные волокна расположены параллельно. Но у ящерицы Tiliqua мышечные веретена, по-видимому, лежат последовательно с экстрафузальными волокнами (Prosser, 1973). Рептилии и амфибии лишены гамма-мотонейронной системы; как интра-, так и экстрафузальные волокна связаны у них с альфа-мотонейронами. У рыб нет мышечных веретен, но в их волокнах находятся рецепторы, чувствительные к их угловой скорости. У членистоногих имеется множество видов рецепторов растяжения; они делятся на два главных типа: 1) лежащие между элементами наружного скелета и реагирующие на вибрации в кутикуле; 2) прикрепленные к сухожилиям и сигнализирующие об изменениях растяжения и давления (Prosser, 1973). Так, крабы обладают рецепторами, которые сигнализируют о положении и движении сустава, а у падальных мух сигналы рецепторов растяжения
в кишке тормозят поедание пищи, когда кишка заполнена. Большинство биологов согласится с тем, что одним из главных эволюционных направлений в животном мире является совершенствование нервной системы: Таким образом, чтобы проследить эволюцию сенсорных процессов, по-видимому, разумно ориентироваться на сложность ее устройства. Однако мы располагаем лишь
очень немногими прямыми данными о нервной системе в прошлом, так как мягкая нервная ткань редко сохраняется в окаменелом состоянии. Косвенные сведения иногда можно получить по остаткам скелета, в особенности по хорошо сохранившемуся черепу позвоночных. Большая часть наших выводов об эволюции нервной системы получена при изучении современных представителей типов животных, о которых известно, что они мало изменились за миллионы лет.
|
||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 857; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.015 с.) |