Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Развитие представлений о сущности жизни. Определение жизни с позиций системного подхода.Содержание книги
Поиск на нашем сайте
БИОЛОГИЯ Развитие представлений о сущности жизни. Определение жизни с позиций системного подхода. Накопленные знания в области биологии и химии во второй половине ХIХ века позволили сделать вывод, что основным субстратом жизни является белок. Ф. Энгельс определил жизнь как «способ существования белковых тел, существенным моментом которого является обмен с окружающей средой. У неорганических тел также может происходить обмен веществ, но разница заключается в том, что обмен неорганических тел разрушает их, а обмен органических тел является необходимым условием их существования». В связи с открытием в 1869 году Мишером нуклеиновых кислот, был пересмотрен и субстрат жизни. Стало ясно, что под субстратом следует понимать комплекс биополимеров – белков и нуклеиновых кислот. В настоящее время не известно ни одной живой системы без совокупности ДНК (или РНК) и белка. Все процессы характеризующие жизнь связаны с комплексными свойствами этих соединений. Главной особенностью субстрата жизни является его упорядоченность на молекулярном уровне (так ДНК представляет собой двухцепочечную молекулу, построенную по принципу комплементарности). Эта упорядоченность приводит, в сою очередь, к формированию надмолекулярных структур (хроматид, хромосом, хромонем, хромомер, фибрилл). Описанная упорядоченность комплекса белка и нуклеиновых кислот в пространстве влечет за собой упорядоченность во времени, что, в конечном итоге, обеспечивает строгую последовательность жизненно важных процессов. Живые системы непрерывно обмениваются с окружающей средой энергией, веществами и информацией, т.е. существуют в форме открытых систем. С потоком вещества и энергии связано самообновление при сохраненных структурах в живом. С потоком информации связана преемственность между сменяющими друг друга биологическими системами – самовоспроизведение и ауторегуляция (саморегуляция), обеспечивающая постоянство структур и внутренней среды – гомеостаз. В связи с изложенным, наиболее точным и современным представляется определение жизни, данное академиком М.В. Волькенштейном: «Живые тела существующие на Земле это есть открытые, саморегулирующиеся, самообновляющиеся, самовоспроизводящиеся системы, состоящие из биополимеров – белков и нуклеиновых кислот».
Строение и виды РНК. Роль различных видов РНК в процессе реализации наследственной информации. См №6 Основные положения хромосомной теории наследственности · Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности: · Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален. · Аллельные гены занимают одинаковые локусы в гомологичных хромосомах. · Гены расположены в хромосоме в линейной последовательности. · Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола). · Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами). · Каждый биологический вид характеризуется определенным набором хромосом — кариотипом. Правила хромосом. Правило постоянства числа хромосом Число хромосом и характерные особенности их строения - видовой признак. Это правилу постоянства числа хромосом. Это число не зависит от высоты организации и не всегда указывает на филогенетическая родство. Например, в ядрах всех клеток лошадиной аскариды Paraascaris megalocephala univalenus находится по 2 хромосомы, в мухи-дрозофилы Drosophila melanogaster - по 8, у человека - по 46, а в речного рака Astacus fluviatalis - по 116. Число хромосом не зависит от высоты организации, а также не всегда указывает на филогенетическое родство: одно и то же число может случаться в очень далеких форм, а в близких видов - очень отличаться. Однако, очень важно, что у представителей одного вида число хромосом в ядрах всех клеток постоянное. Правило четности хромосом Существует также правило четности хромосом, по которому число хромосом всегда является четным, так как в кариотипе хромосомы объединяются в гомологичные пары.
Правило индивидуальности хромосом Гомологичные хромосомы одинаковые по форме и строению, расположением центромер, хромомер, других деталей строения. Негомологическом хромосомы всегда имеют различия. Поэтому имеем правило индивидуальности хромосом: каждая пара гомологичных хромосом характеризуется своими особенностями. Правило непрерывности хромосом Правило непреривности хромосом: в последовательных генерациях число и индивидуальность хромосом сохраняется благодаря способности хромосом к авторепродукции при делении клетки. Репликация ДНК совершается в 2 этапа: 1. сначала цепи полностью раскручиваются и разделяются; 2. затем каждая цепь по отдельности действует как матрица, в результате чего образуются 2 новые двуспиральные молекулы ДНК. Репликация ДНК лежит в основе: воспроизведения генетической информации при размножении живых организмов; передачи наследственных свойств из поколения в поколение; развития многоклеточного организма из зиготы. Общие принципы репликации ДНК применимы, с небольшими модификациями, ко всем организмам. Фрагмент ДНК от одной точки начала репликации до другой точки образует единицу репликации – репликон. Согласованность репликации и клеточного деления у эукариот: Каждый репликон должен реплицироваться один – и только один раз за клеточный цикл. Группы репликонов эукариот (т.н. репликационные единицы, содержащие 20-80 репликонов) инициируются и проходят репликацию упорядоченно, примерно в течение 1 часа. репликация всей ДНК происходит в синтетический период интерфазы клеточного цикла. Спираль родительской ДНК раскручивается и комплементарные цепи отделяются друг от друга. Область расхождения полинуклеотидных цепей в зонах репликации называют репликационными вилками. Поскольку две цепи, образующие молекулу ДНК, соединены водородной связью и переплетены, образуя двойную спираль, их не так-то просто разделить. Более того, ДНК внутри клетки также свернуты плотной сверхспиралью для компактности. Прежде чем разделить цепи, нужно расплести как сверхспираль ДНК внутри клетки, так и двойную спираль ДНК. Это происходит в два этапа. Сначала расплетается сверхспираль ДНК внутри клетки при помощи фермента, известного под названием ДНК-гираза. Как только сверхспираль ДНК внутри клетки расплетена, фермент ДНК-хеликаза раскручивает двойную спираль ДНК. Хеликаза не разрывает цепи ДНК, а просто разрушает водородные связи, которые соединяют цепи молекулы ДНК. Репликационная вилка включает в себя вращающуюся часть, где ДНК раскручивается ДНК-гиразой, затем сразу действует хеликаза, а отрезки одноцепочечной ДНК разъединяются. Для удержания на расстоянии новых цепей имеется специальный «разводящий» белок, который крепится к каждой непарной одноцепочечной ДНК. Этот белок называется SSB -белок. Очень важным моментом в процессе репликации является образование пар оснований: A и T пары и G и C пары. Каждая из разделенных родительских цепей ДНК служит цепью-образцом для синтеза новой комплементарной цепи. Нуклеотиды, пребывающие для формирования новой цепи, определяют своих партнеров, подбирают пару, и таким образом выстраиваются на цепи-образце.
Фермент, который связывает нуклеотиды, известный как ДНК-полимераза также может распознать, правильно ли подобраны пары основаниям. Если неправильно, то неподходящее основание отвергается. ДНК-полимераза состоит из двух субъединиц. Одна из них называется синтетической субъединицей и отвечает за построение новой ДНК. Другая субъединица имеет форму бублика и перемещается вверх или вниз на цепи-образце ДНК, как кольцо на карнизе для штор. Эта субъединица, напоминающая скользящий зажим, скрепляет синтетическую единицу фермента с ДНК. Синтез всегда происходит от 5/-конца к 3/-концу: нуклеотид добавляется к 3/-OH растущей цепи. Хотя ведущая цепь все увеличивается и увеличивается, отстающая цепь сильно запаздывает. После прохождения через репликационную вилку, отстающая цепь представляет собой короткие отрезки с пустотами между ними. Эти короткие, только что созданные отрезки ДНК, названы фрагментами Оказаки (200-400 нуклеотидов) в честь открывшего их ученого. Синтезирует запаздывающую цепь ДНК-полимераза-III малыми фрагментами. Но эти фрагменты нужно соединить, чтобы получить сплошную цепь ДНК. Это соединение берут на себя два фермента, выполняющие свои функции один за другим: ДНК-полимераза I и ДНК-лигаза. ДНК-полимераза-I заполняет собой пустоты, а ДНК-лигаза зашивает одноцепочечный разрыв. Удивительно, что ДНК-полимераза сама не способна индуцировать синтез новых цепей, а может только добавлять звенья к уже имеющейся цепи. Еще более удивительно то, что новые цепи начинают синтез с короткого отрезка, но не ДНК, а РНК! Эти короткие участки РНК называются затравкой, или праймером, а фермент, который начинает синтез новых звеньев построением РНК-затравки называется праймазой. Праймаза индуцирует репликацию всей лидерной цепи, а в отстающей - в каждом фрагменте Оказаки праймаза инициирует репликацию праймера. При каждом строительстве нового фрагмента ДНК, праймаза внедряется и выделяет короткую РНК затравку, чтобы начать процесс синтеза. И только после этого ДНК-полимераза-I начинает наращивать цепь. РНК-затравка-праймер удаляется РНКазой H и 5/ - 3/ экзонуклеазой. После прохождения через репликационную вилку, двуцепочечная молекула ДНК автоматически сворачивается в спираль. Репликация ядерной ДНК происходит в определенный период жизненного цикла клетки – синтетический период интерфазы. Ауторепродукция хромосом и последующее за ней деление клетки направлены на то, чтобы дочерние клетки получили всю полноту генетической информации, имеющуюся в исходной родительской клетке.
Строение сперматозоидов Мужские половые клетки - сперматозоиды (спермии), развивается в очень их несколько тысяч миллионов. Они невелики по размерам (у человека около 70мкм), обладают способностью к активному движению со скоростью 30-50 мкм/сек. Сперматозоид имеет жгутиковую форму. ¨Процесс образования и созревания сперматозоидов - сперматогенез. Сперматозоид состоит из двух частей: 1) головки; 2) хвоста. Головка сперматозоида (caput spermatozoidi) содержит небольшое плотное ядро с гаплоидным набором хромосом. Для человека характерно наличие в ядре 22 аутосомы и 1 половой хромосом (гоносомы). В зависимости от того, какую половую хромосому имеет ядро сперматозоида Х или Y, они делятся на два вида: 1) андроспермии - содержат Y - хромосомы, 2) гинекоспермии - содержат Х - хромосомы. ¨Ядро характеризуется высоким содержанием нуклеопротаминов и нуклеогистонов. Передняя часть ядра покрыта плоским мешочком, который образует чехлик серматозоида. На переднем полюсе чехлика располагается акросома (от греч. acros - верхушка; soma - тело). Оба образования (чехлик и акросома) являются производными комплекса Гольджи. Акросома содержит набор ферментов, среди которых важное место принадлежит гиалуронидазе и протеазам(трипсин), которые способны растворять оболочки яйцеклетки. Головка снаружи покрыта клеточной мембраной. Хвост (feagellum) сперматозоида состоит из: а) связующей части (шейка) образованной двумя центиолями - проксимальной и дистальной, от дистальной берет начало осевая нить (аксонема); б) промежуточной части образованной двумя центральными и 9 парами периферических микротрубочек, окруженных по спирали митохондриями (митохондриальное влагалище); в) главной части, которая по строению напоминает ресничку. Окружена тонкофибриллярным влагалищем; г) терминальной части, которая содержит единичные сократительные филаменты. Также как и головка, хвост покрыт клеточной мембраной Функции сперматозоидов: 1. Оплодотворение яйцеклетки. При помощи хвоста сперматозоид способен двигаться в определенном направлении, которое определяется специфическими веществами, выделяемыми яйцеклеткой - гиногамонами. 2. Реагируют на химические раздражители - хемотаксис. 3. Могут двигаться против тока жидкости - реотаксис. 4. Сохраняют способность к оплодотворению в оптимальных условиях в течение 36-88 часов. 5. Оптимальными условиями является слабощелочная среда.
Транскрипция Процесс переписывания информации о первичной структуре белка с молекулы ДНК на про-и-РНК называется транскрипцией. Синтез про-и-РНК начинается с обнаружения РНК-полимеразой особого участка в молекуле ДНК, который называется промотором - он указывает место начала транскрипции. РНК-полимераза обеспечивает раскручивание участка ДНК, соответствующего транскрибируемому гену, разрушение водородных связей между тяжами ДНК, рождение тяжей, осуществление синтеза про-и-РНК. Сборка рибонуклеотидов в цепь происходит с соблюдением их комплементарности нуклеотидам ДНК. РНК - полимераза способна собирать полинуклеотид от 5' конца к 3' - концу, матрицей для транскрипции может служить только одна из цепей ДНК, та, которая обращена к ферменту своим 3' - концом (3' → 5'). Такую цепь называют кодогенной. Антипараллельное соединение двух полинуклеотидных цепей в молекуле ДНК позволяет РНК полимеразе правильно выбрать матрицу для синтеза про-и-РНК. Транскрипция осуществляется до тех пор, пока РНК-полимераза не встретит специфическую последовательность нуклеотидов - терминатор транскрипции. В этом участке РНК - полимераза отделяется как от ДНК, так и от вновь синтезированной молекулы про-и-РНК. Затем про- и-РНК - специальными ферментами отделяется с ДНК. Образуемая в ходе транскрипции молекула про-и-РНК является точной копией гена и отражает его интрон-экзонную структуру. Тройки рядом стоящих нуклеотидов, шифрующие аминокислоты называют кодонами. На специальных генах синтезируются и два других вида РНК: т-РНК и р-РНК. Начало и конец синтеза всех типов PНK на матрице ДНК строго фиксирован специальными триплетами, которые инициируют запуск и остановку (терминацию) синтеза. Процессинг - это созревание и-РНК. Происходит удаление из первичных транскриптов неинформативных для данного блока интронных участков, размер которых варьирует от 100 до 1000 нуклеотидов. На долю интронов приходиться около 80 % всей про-и-РНК. Удаление интронов с последующим соединением экзонных участков называют сплайсингом. Закономерность вырезания интронов обеспечивается благодаря налипанию на их концы специфических нуклеотидных последовательностей, опознаваемых определенными ферментами. После сшивания смысловой части и-РНК осуществляется ориентация ее концов: на 5' – конце происходит метилирование азотистых оснований, образуется колпачок – КЭП, обеспечивающий узнавание молекул и-РНК малыми субчастицами рибосом. На 3' - конце первичного транскрипта присоединяется последовательность, состоящая из 100 - 200 остатков адениловой кислоты (поли А). Поли А определяет кратность трансляции. К тому же эта последовательность способствует выходу зрелой м-РНК из ядра.
Благодаря преобразованиям, происходящим с про-и-PНK в ходе процессинга, зрелая и-РНК эукариот характеризуется большой стабильностью. После завершения процессинга зрелая иРНК проходит отбор перед выходом в цитоплазму, куда попадает всего 5% иРНК. Остальная часть расщепляется не выходя из ядра. 2) Трансляция - это процесс считывания наследственной информация с последовательности нуклеотидов иРНК на последовательность аминокислот в полипептидной цепи. Процесс обеспечивается взаимодействием тРНК и иРНК. Осуществляется на рибосомах. В рибосомах имеется две бороздки: одна удерживает растущую полипептидную цепь, другая – и-РНК. Кроме того в рибосомах имеются два участка, связывающих тРНК. В аминоацильном участке (А - участке) размещается аминоацил тРНК, несущая определенную аминокислоту. В пептидильном участке (П - участок) располагается обычно тРНК, которая нагружена цепочкой аминокислот, соединенных пептидными связями. Образование А и П участков обеспечивается обеими субчастицами рибосомы. При реализации генетической информации каждая тРНК распознает, присоединяет и переносит в рибосому свою аминокислоту. Этот процесс называется рекогниция. Специфическое соединение тРНК со своей аминокислотой протекает в два этапа и приводит к образованию соединения, называемого аминоацил - тРНК. Процесс этот происходит при участии специфического фермента (аминоацил - тРНК синтетазы). В ходе трансляции можно выделить 3 фазы: инициацию, элонгацию и терминацию. 3) Фаза инициации, или начало синтеза пептида. Заключается в объединении большой и малой субчастиц рибосомы на определенном участке и-РНК и присоединении к ней первой аминоацил тРНК. В молекуле любой и-РНК вблизи ее 5' - конца имеется участок, комплементарный р-РНК малой субчастицы рибосомы и специфически узнаваемый ею. Рядом с ним располагается стартовый кодон (инициирующий) АУГ, шифрующий аминокислоту метионин. Малая субъединица рибосомы соединяется с иРНК таким образом, что стартовый кодон АУГ располагается в области, соответствующей П - участку. При этом только инициирующая тPHK, несущая метионин способна занять место в недостроенном П - участке малой субчастицы рибосомы и комплементарно соединиться со стартовым кодоном. После этого происходит объединение большой и малой субчастиц рибосомы с образованием ее пептидильного и аминоациального участков. К концу фазы инициации П участок занят аминоацил-тРНК, связанной с метионином, а в А-участке рибосомы располагается следующий за стартовым кодон. Процессы инициации, трансляции катализируются особыми белками - факторами инициации, которые подвижно связаны с малой субчастицей рибосомы. 4) Фаза элонгации, или удлинения пептида. Включает в себя реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил-тРНК очередного кодона, находящегося в А - участке, комплементарное взаимодействие между кодоном и антикодоном. Благодаря особенностям строения тРНК при соединении ее антикодона с кодоном и-РНК, транспортируемая ею аминокислота, располагается в А - участке поблизости от ранее включенной аминокислоты, находящейся в П – участке. Здесь между аминокислотами образуется пептидная связь, катализируемая особыми белками, входящими в состав рибосомы В результате предыдущая аминокислота теряет связь со своей т-РНК и присоединяется к аминоацил-т-РНК, расположенной в А - участке. Находившаяся в этот момент в П – участке тРНК высвобождается и уходит в цитоплазму. Перемещение т-РНК, нагруженной пептидной цепочкой из А - участка в П участок сопровождаемся продвижением рибосомы по и-РНК на шаг, соответствующий одному кодону. Затем следующий кодон входит в контакт с А - участком, где он будет специфически «опознан» соответствующей аминоацил-тРНК, которая разместит здесь свою аминокислоту. Такая последовательность событий повторяется до тех пор, пока в А - участок рибосомы не поступит кодон - терминатор, для которого не существует соответствующей т-РНК. Скорость элонгации зависит от различных факторов, в том числе и от t°. Современная теория гена. Ген (совр.) – единица генетического материала, передающаяся от родителей к потомству, которую возможно обнаружить в эксперименте по е способности мутировать в различные состояния, рекомбинировать с другими подобными единицами и функционировать, наделяя фенотипом. Ген (программа «Геном человека») – единица транскрипции, которая может быть транслирована в одну или несколько аминокислотных последовательностей.
Оценка числа генов у человека составляет величину 60 000 – 70 000, из них - «гены домашнего хозяйства» 14 000, транскрибируемая часть генома - 20 000.
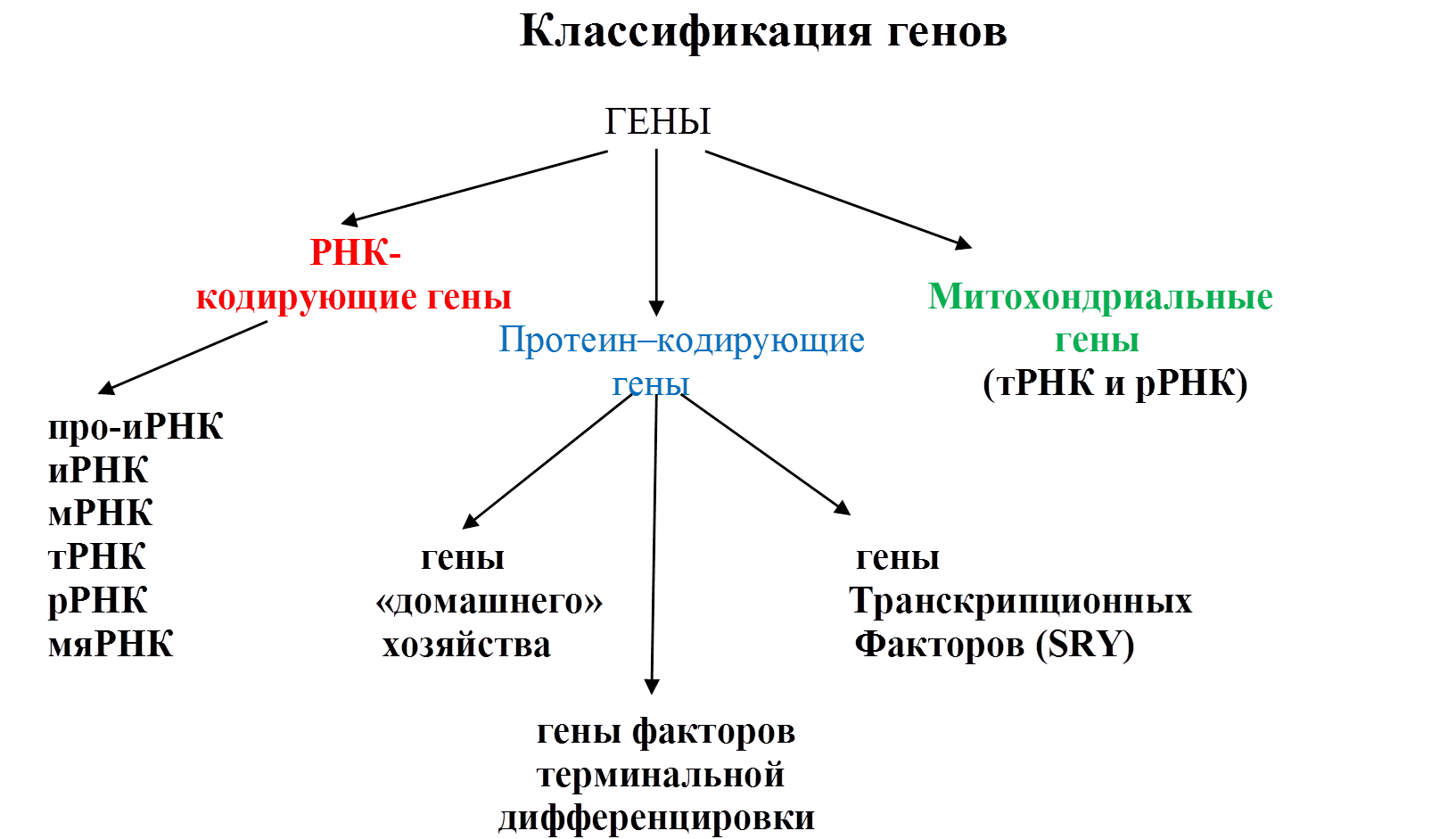
РНК-кодирующие гены определяют синтез РНК, необходимый для обеспечения процессов сплайсинга, синтеза рибосом и процессов трансляции. РНК-кодирующие гены дают информацию для синтеза молекул РНК (тРНК, рРНК, иРНК), обладающих регуляторным действием, т.е. влияющих на функции других генов. Например, РНК для выключения или инактивации одной из X-хромосом у женщин. Гены, кодирующие малые ядерные РНК: малые ядерные РНК помогают удалять интроны из проматричной РНК. Этот процесс осуществляется таким образом, что следующие друг за другом экзоны, т.е. кодирующие фрагменты мРНК, никогда физически не разобщаются. Экзоны соединяются между собой с помощью молекул так называемых малых ядерных РНК. Молекулы малых ядерных РНК играют роль временных матриц, удерживающих близко друг от друга концы двух экзонов, для того чтобы сплайсинг произошел в правильном месте. После того, как таким путем из про-иРНК удаляются все интроны, завершается процессинг, мРНК и зрелая рРНК покидают ядро. Гены, кодирующие белки (протеин-кодирующие), по своей структуре и функциям разделяются на гены «домашнего хозяйства» или гены жизнеобеспечения клетки. Имеются также гены специальных функций, т.е. гены терминальной дифференцировки. Они кодируют белки, характерные для дифференцируемой ткани и определяющие её основные функции. В последнее время выделяют гены особых ядерных белков, названных транскрипционными факторами. Имея сравнительно небольшие размеры, эти гены характеризуются наличием высококонсервативных последовательностей, белковые продукты которых способны соединяться с регуляторными областями ДНК многих структурных генов, вызывая их репрессию или активацию (SRY). Митохондриальные гены: примерно 95 % ДНК находятся в спирализованном состоянии в ядре каждой клетки организма и только около 5% сосредоточены в многочисленных митохондриях (около 1000 на одну клетку), двумембранных органоидах, отвечающих за дыхание и энергетические процессы. Митохондриальные ДНК содержат всего 37 генов. 13 генов кодируют полипептиды, входящие в состав комплекса (NADH-дегидрогеназы-коэнзим Q оксидоредуктазы). Остальные 24 гена необходимы для трансляции белка на митохондриальных рибосомах. Генетический код митохондрий несколько отличается от ядерного, поэтому синтез белка в них требует присутствия собственного вспомогательного аппарата. Из этих 24 генов 22 гена кодируют тРНК, а 2 гена – рРНК митохондрий. Этапы реализации наследственной информации. Трансляция и посттрансляционные процессы. Механизм осуществления. Условия трансляции. Значение качественного и количественного состояния трансляционных ультраструктур. Реализация генетической информации, закодированной в ядерной ДНК, её расшифровка происходит в результате синтеза полипептидных цепей. Таким образом, реализация наследственной информации в клетке осуществляется в два этапа: сначала информация о структуре белка копируется с ДНК на иРНК (транскрипция), а затем реализуется на рибосоме в виде конечного продукта — белка (трансляция). I. Т ранскрипция – процесс перезаписи генетической нформации из ДНК в РНКовый текст. Транскрипция, или переписывание, происходит не на всей молекуле ДНК, а на участке, отвечающем за определенный белок (ген). Условия, необходимые для транскрипции: а) раскручивание участка ДНК с помощью расплетающих белков - ферментов б) наличие строительного материала в виде АТФ. ГТФ. УТФ. 1ДТФ в) ферменты транскрипции - РНК-полимеразы I, II, III г) энергия в виде АТФ. Транскрипция происходит по принципу комплементарности. При этом с помощью специальных белков-ферментов участок двойной спирали ДНК раскручивается, является матрицей для синтеза иРНК. Далее вдоль цепи ДНК движется фермент РНК-полимераза, соединяя между собой нуклеотиды по принципу комплементарности в растущую цепь РНК. Далее одноцепочечная РНК отделяется от ДНК и через поры в мембране ядра покидает клеточное ядро. II. Трансляция. Суть трансляции -перевод 4-хбуквенного языка мРНК на 20-тибуквенный язык белка. Процесс трансляции состоит в переносе закодированной в иРНК генетический информации в аминокислотную последовательность белка. Осуществляется биосинтез белка в цитоплазме на рибосомах и состоит из нескольких этапов: 1. Подготовительный этап (активация аминокислот): состоит в ферментативном связывании каждой аминокислоты со своей тРНК и образовании комплекса аминокислота - тРНК. 2. Собственно синтез белка, который включает три стадии: а) инициация - иРНК связывается с малой субъединицей рибосомы, первыми кодонами, инициирующими, являются АУТ или ГУГ. Этим кодонам соответствует комплекс метионил -тРНК. Вместе с тем, в инициации участвует три белковых: фактора, облегчающие связывание мРНК с большой субчастицей рибосомы, образуется инициаторный комплекс б) элонгация - удлинение полипептидной цепочки. Процесс осуществляется в 3 шага и состоит в связывании кодона мРНК с антикодоном тРНК по принципу комплементарности в активном центре рибосомы, затем в образовании пептидной связи между двумя остатками аминокислот и перемещении дипептида на шаг вперёд и, соответственно, передвижения рибосомы вдоль иРНК на один ко дон вперед в) терминация - окончание трансляции, зависит от присутствия в иРНК терминирующих кодонов или "стоп-сигналов" (УАА,УГА,УАГ) и белковых ферментов - факторов терминации. В клетке для синтеза белка используется не одна, а несколько рибосом. Такой работающий комплекс иРНК с несколькими рибосомами принято называть полирибосомой. В таком случае синтез белка происходит быстрее, чем при использовании только одной рибосомы. Уже в ходе трансляции белок начинает укладываться в трёхмерную структуру, а при крайне важно сти в цитоплазме принимает четвертичную организацию. III. Посттрансляционные процессы Многие ферменты после синтеза белковой цепи претерпевают модификации, без которых фермент не проявляет свою активность в полной мере. Такие модификации называются посттрансляционными модификациями (процессингом). Один из самых распространенных типов модификации — присоединение химических групп к боковым остаткам полипептидной цепи. Например, присоединение остатка фосфорной кислоты называется фосфорилированием, оно катализируется ферментом киназой. Многие ферменты эукариот гликозилированы, то есть модифицированы олигомерами углеводной природы. Ещё один распространенный тип посттранляционных модификаций — расщепление полипептидной цепи. Например, химотрипсин (протеаза, участвующая в пищеварении), получается при выщеплении полипептидного участка из химотрипсиногена. Химотрипсиноген является неактивным предшественником химотрипсина и синтезируется в поджелудочной железе. Неактивная форма транспортируется в желудок, где превращается в химотрипсин. Такой механизм необходим для того, чтобы избежать расщепления поджелудочной железы и других тканей до поступления фермента в желудок. Неактивный предшественник фермента называют также «зимогеном». Рибосома ИРНК, встроенная в рибосому Причины мутаций Установлено, что появление мутаций детерминируется изменениями в молекулах ДНК, возникающими на основе нарушений в метаболизме организма и под прямым влиянием факторов среды. Эти принципы современной теории мутаций показывает всю опасность загрязнения среды мутагенами. Причины появления мутаций могут корениться в естественных условиях среды, в особенностях обмена веществ. Мутации, возникающие от этих причин, получили название естественных, или спонтанных, мутаций. В наше время обнаружено много факторов, с помощью которых мутации могут быть вызваны искусственно. Мощным мутагеном является действие разных видов радиации, химические мутагены и другое. Мутации, получаемые под их воздействием, называют индуцированными мутациями. Факторы, вызывающие мутации, называются мутагенами. Факторы окружающей среды называются экзомутагены, а продукты метаболизма в организме, способные вызвать мутации, называются эндомутагенами. Экзомутагены поступают в организм человека из атмосферы, с водой, пищей, с лекарственными препаратами. Экзомутагены классифицируются на физические, химические и биологические. К физическим мутагенам относятся все виды ионизирующего излучения (электромагнитные и корпускулярные). К более слабым физическим мутагенам относятся колебания to, давления, ультразвук, инфразвук, вибрация, шум (производственный). Химические мутагены характеризуются большим многообразием и имеют природное и антропогенное происхождение. Особые опасения вызывают искусственно синтезированные химические соединения – ксенобиотики различной природы: микотоксинов, нитрозосоединений, растительных алкалоидов, гетероциклических аминов, флавоноидов, отдельных ароматических углеводородов и т.д. К биологическим мутагенам относятся вирусы, простейшие, бактерии, гельминты. Вирусы - «живые» мутагены, они приносят в клетки собственную генетическую информацию, изменяя хромосомный аппарат (онкогенные вирусы, ретровирусы оспы, гриппа, коревой краснухи). Ретровирусы например вызывают лейкозы у людей, увеличивают число аутоиммунных болезней (эритематозная волчанка, аутоиммунный гломерулонефрит).
Генетический полиморфизм.
Полиморфизм – отличия последовательности ДНК в геноме у представителей одного вида. В отличие от мутаций, приводящих к патологическим изменениям и снижающих жизнеспособность, генетические полиморфизмы проявляются менее очевидно. Далеко не всегда они являются нейтральными, чаще они приводят к появлению измененных белковых продуктов и их функциональной активности. Так, однонуклеотидные замены (SNP) в экзонах гена влияет на изменение третичной структуры белка, пострансляционную модификацию. При этом функциональный спектр таких белков сильно меняется от практически нейтрального эффекта до полного нарушения функции белкового продукта. В определенных условиях некоторые генетические полиморфизмы могут предрасполагать, либо, наоборот, препятствовать проявлению различных заболеваний. Гены, аллельные варианты которых при наличии определённых условий предрасполагают к определенным заболеваниям называются генами предрасположенности. Таким образом, гены предрасположенности - это мутантные гены, которые совместимы с рождением и жизнью в постнатальном периоде, но при определенных неблагоприятных условиях способствуют развитию того или иного заболевания (кардиомиопатии, атеросклерозу, эссенциальной гипертензии, бронхиальной астме, ИБС, сахарного диабета типа I, II, поликистоза почек, псориаза, эпилепсии.
Геномный импринтинг. Механизмы геномного импринтинга. Однородительская дисомия (ОРД). Болезни импринтинга. Млекопитающие являются диплоидными организмами, чьи клетки обладают двумя совмещенными наборами хромосом, один из которых унаследован от матери, а другой - от отца. Таким образом, млекопитающие обладают двумя копиями каждого гена. В норме и материнская, и отцовская копии каждого гена имеют одну и ту же возможность быть активной в любой клетке. Геномный импринтинг - это эпигенетический механизм, который изменяет эту потенцию, потому что он ограничивает экспрессию гена одной из двух родительских хромосом. Это явление демонстрируют лишь несколько сотен из примерно 25000 генов в нашем геноме, а большинство экспрессируются в равной степени, будучи унаследованы от любого из родителей. Определение геномного импринтинга ограничено здесь "специфичной, в отношении родителя, экспрессией генов в диплоидных клетках". Таким образом, диплоидные клетки, содержащие по две родительские копии всех генов, будут экспрессировать только одну родительскую копию импринтированного гена и сайленсировать другую родительскую копию. Напротив, неимпринтированные гены будут экспрессироваться в диплоидной клетке обеими родительскими копиями. Для понимания концепции импринтированных генов важно различать импринтированные гены и гены, демонстрирующие кажущуюся специфичную в отношении родителя экспрессию из-за неравного родительского генетического вклада в зародыш. Примерами неравного родительского генетического вклада являются гены, сцепленные с Y-хромосомой и присутствующие только у самцов, гены, избежавшие Х-инактивации у самок, митохондриальные гены, привносимые в основном матерью, и иРНК и белки, присутствующие в цитоплазме только спермия или только яйцеклетки. Однородительская дисомия, то есть наследование обеих копий целой хромосомы или ее части от одного родителя (при отсутствии соответствующего генетического материала от другого родителя), является исключением из менделевских принципов наследования. Она встречается редко и вызывает, например, синдром Прадера-Вилли и синдром Ангельмана. Роль дисомии в патологии во многом усугубляется геномным импринтингом, который приводит к неодинаковой экспрессии материнской и отцовской копий гена. Возможный механизм дисомии - элиминация лишней хромосомы у плода с трисомией на ранних стадиях эмбриогенеза. Болезнь проявляется в том случае, если элиминируется лишняя хромосома, происходящая из нормальной гаметы. Однородительская дисомия была описана при муковисцидозе, когда оба мутантных аллеля наследовались от одного родителя. В таких случаях дисомия имитирует аутосомно-рецессивное наследование. У 20-30% больных с синдромом Прадера-Вилли, имеющих по данным цитогенетического исследования нормальный кариотип, с помощью молекулярно-биологических методов обнаруживается дисомия материнской 15-й хромосомы. Отцовская 15-я хромосома у таких больных отсутствует. К настоящему времени идентифицировано несколько механизмов возникновения БГИ. 1. Однородительская дисомия (ОРД) — наличие у больного двух хромосом с импринтинговыми участками, полученными от одного из родителей. В этом случае человек имеет нормальный кариотип (46 хромосом), однако, обе хромосомы одной из пар получены им от одного из родителей. Причин возникновения ОРД несколько, но наиболее вероятны из них четыре: а) нерасхождение хромосом во втором мейотическом делении с образованием совершенно идентичных хроматид; б) нерасхождение хромосом в первом делении мейоза с возникновением негомологичных хромосом; в) исправление возникшей при делении клеток трисомии или моносомии по хромосомам, содержащим импринтированные гены; г) соматическая рекомбинация хром
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-04-04; просмотров: 151; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.21.105.46 (0.018 с.) |