
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Психофизиология памяти в школе Е.Н. СоколоваСодержание книги
Поиск на нашем сайте Объективное изучение ориентировочной реакции в контексте памяти было начато Е.Н. Соколовым и его коллегами в 50—60-е гг. Впервые в психологии в опытах использовались методы регистрации объективных показателей физиологических функций. Анализировали электроэнцефалограмму (ЭЭГ), частоту биения сердца, частоту дыхания, движения глаз, сопротивление кожи (КГР), плетизмограмму. Основной эксперимент был задуман так, что получали массу возможностей для исследования привыкания — пластичности, играющей важнейшую роль в ориентировочно-исследовательском поведении. Привыкание — негативная форма обучения, обнаруживаемая у всех живых существ, независимо от уровня их эволюционного развития. Привыкание стало тем феноменом, который положил начало систематическому исследованию механизмов памяти. Анализировались изменения, происходящие на макроуровне. В экспериментах Е.Н. Соколова была исследована динамика привыкания, выявлены его основные критерии, получены электрофизиологические показатели. Однако по мере накопления экспериментальных данных и появления новых научных гипотез выбор объектов для исследований расширялся — начавшись с исследований привыкания на уровне человека, в настоящее время он включает анализ нейрогенетических механизмов, изучение генетического контроля эффективности работы синаптических контактов во время формирования пластических изменений. Исследование памяти методом угашения ориентировочного реф лекса. Каждый новый раздражитель, в том числе световой, вызывает комплекс соматических, вегетативных и электроэнцефалографических реакций — генерализованную ориентировочную реакцию. Но по мере повторения светового сигнала генерализованный ориентировочный рефлекс сменяется изменениями, ограниченными зрительной системой — локальным ориентировочным рефлексом, угасание которого развивается при дальнейшем применении раздражителя. Наиболее характерным выражением локального ориентировочного рефлек- са является депрессия альфа-ритма затылочной области при действии Реакции депрессии альфа-ритма характеризуются определенным латентным периодом и длительностью. Фоновая ЭЭГ человека, находящегося в темноте с закрытыми глазами, выражена высокоамплитудным альфа-ритмом, лишь на короткое время сменяющимся депрессией. Наиболее вероятна в этих условиях депрессия длительностью 1—2 с. При открывании глаз число спонтанных реакций возрастает. Появление в поле зрения постоянной световой точки приводит к тому, что число реакций депрессии альфа-ритма, прежде всего тонических, еще более увеличивается. Однако следствием длительной фиксации светящейся точки является постепенное (2—12 опытов) развитие привыкания к ней. В результате спонтанные реакции депрессии альфа-ритма тонического типа исчезают, частота фазических реакций снижается, и ЭЭГ возвращается к типу, характерному для ЭЭГ при открытых глазах испытуемого в отсутствие фиксации. Что касается реакций депрессии альфа-ритма на световые раздражители разной интенсивности, то они в условиях непрерывной смены уровней интенсивности при применении 50—60 стимулов в опыте не угасают. Параметры депрессии альфа-ритма и вероятность попаданий закономерно связаны с интенсивностью раздражителя. В условиях нанесения разных по силе раздражителей с усилением раздражителя закономерно сокращается среднее значение и дисперсия латентного периода, возрастает вероятность попаданий. Длительность реакции депрессии альфа-ритма максимальна в области порогового значения стимула. Можно предположить, что подобная форма зависимости длительности реакции ЭЭГ от интенсивности стимула возникает в силу того, что реакция депрессии альфа-ритма в условиях предъявления сигналов разной интенсивности зависит от двух факторов: физической силы раздражителя и трудности обнаружения сигнала при приближении к порогу. Как меняются рассмотренные параметры реакций депрессии альфа-ритма в ходе многократных применений раздражителей разной интенсивности? Опыты показали, что у большинства испытуемых угашение наступает после 300—500 раздражений. В тех случаях, когда угашение развивалось, оно затрагивало либо одновременно все интенсивности раздражителей, либо могло начинаться с любой из них. При угашении сначала происходит сокращение длительности депрессии альфа-ритма и лишь потом снижается вероятность попаданий. При этом величина латентных периодов практически не меняется и определяется лишь интенсивностью стимула. Наблюдающееся в ряде случаев увеличение латентного периода связано с формальным примене- 5-1015 66 Нейрофизиология Клеточные основы обучения 7 Механизмы научения 67
Из приведенных опытов следует важный вывод о том, что латентный период депрессии альфа-ритма определяется прежде всего физической силон раздражителя, тогда как длительность реакции и вероятность ее появления зависит не только от физической силы, но и от степени достигнутого угасания. Более того, в области надпороговых раздражителей длительность депрессии альфа-ритма и вероятность попадания определяется прежде всего степенью угашения реакции. Угашение ориентировочного рефлекса состоит в потенциации синапсов вставочных нейронов. При этом афферентное коллатеральное торможение усиливается настолько, что реакция на выходе нейрона блокируется. Потенциированная система синапсов, соответствуя системе признаков повторяющегося сигнала, обеспечивает избирательность угашения ориентировочного рефлекса в отношении свойств наносимого раздражителя. При изменении раздражителя возбуждение поступает через новые синапсы, для которых афферентное коллатеральное торможение не выработано. Реакция на новый раздражитель тем больше, чем больше включено новых синапсов, т.е. чем большим количеством свойств новый раздражитель отличается от применявшегося ранее в опыте. Гипотеза о том, что каждому афферентному входу соответствует особый вставочный нейрон, предполагает чрезвычайно большое число вставочных нейронов. Это приводит к предположению о том, что каждому синапсу на пирамидном нейроне соответствует образованный коллатерально синапс на вставочном нейроне. При этом потенциация, ведущая к возрастанию параллельного торможения, возникает в возбуждающих синапсах на теле вставочного нейрона. Повторение раздражителя приводит к сокращению латентного периода тормозного эффекта в отношении многократно применяемого в опыте стимула. Другой раздражитель, адресованный к другому синаптическому аппарату того же вставочного нейрона, но не «подготовленного» постси-наптической потенциацией, окажется неэффективным, вставочный нейрон не возбудится и не вызовет торможения. Пирамидный нейрон не будет заторможен и прореагирует. Характерной чертой угашения ориентировочного рефлекса является высокая избирательность в отношении параметров стимула. Представление о синаптическом механизме «привыкания» хорошо согласуется с этими данными. Известно, что постсинаптическая потенциация избирательно возникает только в тех синапсах, которые непосредственно участвуют в проведении сигналов. Таким образом, на уровне нейрона «внимания» следует ожидать избирательного угашения реакций на сигнал, специфически связанный с той, системой синапсов, которая участвует в кодировании наносимого раздражителя. Специально следует остановиться на эффекте растормаживания, который позволяет отличать эффект ослабления реакции от ее угашения. После действия нового раздражителя реакция на прежний стимул временно восстанавливается. В рамках рассмотренной схемы этому соответствует временное выключение вставочных нейронов в цепи параллельного торможения. Это можно объяснить, предположив, что кроме возбуждающих синапсов на вставочных нейронах имеются тормозящие синапсы с высоким порогом, через которые осуществляется влияние детекторов новизны. При сильном возбуждении детекторов новизны вставочные нейроны временно отключаются, вызывая эффект растормаживания. На основании имеющихся данных можно высказать предположение о том, что детекторы новизны расположены на разных уровнях ЦНС. Наиболее яркое участие в реакции новизны принимают нейроны гиппокампа и глубоких слоев коры. Что касается угасания реакций неспецифического таламуса и неспецифической системы среднего мозга, то не исключено, что они получают готовый сигнал новизны и поэтому угашение реакции у них отражает исчезновение реакции на новизну на другом уровне. Определение конфигурации следа. Определение конфигурации следа памяти методом регистрации ориентировочного рефлекса основано на том, что его компоненты избирательно угасают в отношении параметров повторяющего сигнала. Изменение любого параметра стимула приводит к появлению ориентировочной реакции, величина которой в известных пределах пропорциональна разнице между наносимым раздражителем и сформировавшимся следом. После угашения реакции, достигнутого в отношении одного раздражителя, систематически изменяя тест-стимулы, можно определить границы сформировавшегося следа. Возможность изучения конфигурации следа памяти путем последовательных проб обусловлена структурой ориентировочного рефлекса, возбуждаемого нервными импульсами, которые возникают при сравнении внешнего сигнала со следом памяти. Систематическое изучение следовых процессов в зрительном анализаторе человека на основе угасания локального ориентировочного ответа в определенной области было проведено в ряде исследований. Задача состояла в том, что, получив угашение реакции депрессии ЭЭГ затылочной области на световой стимул фиксированной интенсивности, цвета, положения в пространстве, длительности и периодичности появления, последовательно изменять разные параметры стимула так, 5* Глава I Пеирофичиолошя Клеточные основы обучения 7 Механизмы научения 69
В ходе опыта непрерывно регистрировалась ЭЭГ затылочной области. После того, как реакции в виде депрессии альфа-ритма угасали, один из параметров сигнала изменялся. Появление реакции при изменении стимула говорило о том, что нервная система отличает тест-стимул от следа стимула-эталона. После однократной пробы, чтобы исключить растормаживающее влияние тест-стимула, несколько раз наносился стимул-эталон. Когда реакция на стимул-эталон вновь устойчиво угасала, изменялся другой параметр сигнала. Были проведены опыты с изменением цвета и пространственного положения стимула'. В этом случае один глаз фиксировал центр периметра, а другой — закрывался повязкой. Тест-стимулы последовательно наносились на соседние участки сетчатки. При обработке данных определялась зависимость длительности ориентировочной реакции или частоты ее появления от изменения тест-стимула относительно изучаемого параметра стимул-эталона. Поскольку ранее избирательное угасание было продемонстрировано в отношении высоты, интенсивности и других параметров звуковых раздражителей, то, обобщая полученные данные, можно сказать, что при многократном повторении раздражителя в памяти с большой точностью фиксируются все параметры сигнала. Таким образом, нервная система, фиксируя в изменении своих элементов совокупность свойств раздражителя, создает в известном смысле «нервную модель стимула», которую можно представить как многомерный самонастраивающийся фильтр, избирательно подавляющий повторяющиеся в опыте сигналы. С этой точки зрения, при одновременном изменении ряда параметров сигнала реакция фильтра определяется суммарным эффектом несовпадения тест-стимула со стимул-эталоном. Опыты показали, что наиболее сильные реакции, включающие ряд вегетативных и соматических реакций, вызывал ритмический раздражитель, где количество измененных параметров было наибольшим. Полученные данные говорят о том, что отдельные признаки сигнала фиксируются самостоятельно и сличение внешнего сигнала со следом происходит по совокупности признаков. 7.2.2. Нервная модель стимула Идея о нервной модели стимула, которая составила в дальнейшем теоретическую основу исследований Е.Н. Соколова и его коллег, возникла в процессе исследования ориентировочной реакции (1969). Основное содержание гипотезы о нервной модели стимула следующее — в нервной системе создается след памяти о качествах действующего стимула. Все параметры фиксируются но мере предъявления стимула. При изменении физических характеристик стимул воспринимается как незнакомый. Действие нового стимула вызывает сигнал рассогласования с нервной моделью стимула, что ведет к растормаживанию реакции. Формирование нервной модели стимула начинается с выделения отдельных признаков сигнала. Это осуществляется сетями, составленными из специфических афферентных нейронов. При этом отдельные признаки сигнала кодируются порядковым номером нейрона, стоящего на выходе нейронной сети. Под влиянием многократных повторений раздражителя афферентные нейронные сети перестраиваются и изменяют свои параметры так, что многократно повторяющиеся признаки сигнала подчеркиваются, т.е. в определенных нейронах эти признаки приобретают большую скорость поведения при осуществлении специфических рефлексов. Другие нейроны ослабляют свое действие. Облегчение одних и инактивация других элементов нейронной сети приводит к стабилизации реакции и позволяет воспринимать знакомые предметы быстрее и определеннее, чем малознакомые объекты. На ряде пирамидных нейронов новой коры и гиппокампа (нейронах «внимания») конвергирует множество признаков внешней среды. При этом, по-видимому, каждой отдельной градации признака сигнала соответствует отдельный синаптический контакт. Время действия раздражителя и интервал между отдельными раздражителями с этой точки зрения кодируется аналогичным образом — номерами нейронов, избирательно реагирующих на интервал между раздражителями. Сигнал поступает на нейрон «внимания» прямым путем в виде возбуждающего постсинаптического потенциала (ВПСП) и параллельно через систему вставочных нейронов, заканчивающихся на нейроне «внимания» тормозящими синапсами в виде тормозного постсинаптического потенциала (ТПСП). В ходе многократного применения раздражителя происходит потенциация синаптических контактов вставочных нейронов, которые были вовлечены в реакцию. Матрица по-тенциированных синапсов, связанных с определенными свойствами раздражителя, хранит «конфигурацию» сигнала и является «нервной моделью стимула». Исследование динамики ориентировочного рефлекса у человека и животных показало, что ориентировочная реакция возникает не на сам стимул, а в результате сличения стимула со следом, оставленным в нервной системе предшествующими раздражителями. Если данный стимул и след, оставленный ранее предъявленными раздражителями, совпадают, то ориентировочная реакция не возни- 70 'laud 1 Нейрофизиология Клеточные основы обучения 7 Механизмы научения 71
Нервная модель стимула как селективный фильтр. Раздражитель, параметры которого строго фиксированы, при первом применении вызывает у человека и животных целый комплекс реакций, включающий движение глаз в направлении стимула, кожно-гальваничес-кую реакцию, сосудистые изменения и депрессию альфа-ритма. Рассмотрим случай, когда раздражителем служит световое пятно, подаваемое с помощью проекционного периметра в определенный участок поля зрения человека. Размер пятна, его интенсивность, длительность и интервал между отдельными предъявлениями строго постоянны. После 10—15 предъявлений стимула компоненты генерализованного ориентировочного рефлекса угасают. Более устойчиво удерживаются локальные проявления ориентировочного рефлекса в виде депрессии альфа-ритма в затылочной области. После 30—40 предъявлений и эта реакция заметно редуцируется или полностью угасает. Тогда один из параметров стимула изменяется. При изменении любого из фиксированных параметров стимула на величину, превосходящую разностный порог, возникает реакция депрессии альфа-ритма, которая иногда сопровождается кожно-гальванической реакцией и движением глаз. Таким образом, под влиянием повторения стимула, обладающего фиксированными параметрами, нервная система селективно блокирует все сигналы, которые лежат в окрестности его фиксированного параметра. Вместе с тем все сигналы, лежащие вне фиксированного параметра, вызывают реакцию. Можно сказать, что нервная система вырабатывает селективный фильтр. Этот фильтр является самонастраивающимся и многомерным. Нервная модель стимула выполняет в таком фильтре функцию маски, отбирающей сигналы. Поиск локализации нервной модели привел к разворачиванию работ на нейронном уровне. Где и в какой форме происходит фиксация параметров действующего стимула? Сначала эксперименты проводили на кроликах, регистрируя электрическую активность нейронов различных структур мозга экстраклеточными микроэлектродами, а затем, по мере углубления представлений о механизмах, участвующих в фиксации характеристик поступающего сигнала, стали выпол- нять опыты на моллюске Limnaea stagnalis и на моллюске Helix рота- tia. Ответ на вопрос о конкретных нейрофизиологических механизмах, участвующих в «запоминании» параметров действующего стимула, искали при помощи микроэлектродной регистрации внутриклеточных процессов, которые развивались в отдельных клетках во время привыкания, а затем и при более сложных формах обучения (Соколов, 1969, 1981). 7.2.3. Простые системы в исследованиях механизмов обучения Сложность признаков сигналов, используемых ЦНС позвоночных животных, затрудняет детальный анализ формирования нервной модели стимула в процессе негативного научения. Переход на нейроны моллюсков позволил изучить этот механизм в более простых условиях. Кроме того, наличие гигантских нейронов в ганглиях моллюсков позволяет осуществить внутриклеточное изучение негативной формы обучения в течение достаточно длительного времени. Поведенческие опыты на улитках показали, что реакция в виде сокращения щупалец по мере повторения раздражения исчезает. Это привыкание специфично в отношении наносимого раздражения и относительно устойчиво, отличаясь от утомления и сенсорного утомления. Так, у Б. Холмгрена и С. Френка (1964) эффект привыкания был продемонстрирован на улитке методом внутриклеточной регистрации реакций отдельных нейронов. На нейронах абдоминального ганглия аплизии было обнаружено привыкание к механическим раздражениям кожи. Исследователи сразу оценили ряд преимуществ, характерных для электрофизиологического исследования гигантских нейронов. Е.Н. Соколов и В.П. Дуленко (1968) в опытах на виноградной улитке изучали привыкание отдельных нейронов. Были использованы те же приемы, что и при изучении «негативного научения» на нервных клетках мозга кролика. В качестве раздражения применяли стимулы, наносимые на кожу ноги. Длящееся прикосновение вызывало эффект лишь своей начальной фазой, при продолжающем действии тактильного раздражителя на одну точку кожи реакция постепенно исчезала. До опытов Е.Н. Соколова и его коллег в 60—70-е гг. никто серьезно не изучал пластические свойства пейсмекерного потенциала нейрона, хотя многих исследователей привлекала эта необычная по своему ге-незу и функциям активность нервных клеток. Моделируя ситуацию привыкания на одной клетке при внутриклеточной регистрации ее электрической активности, Е.Н. Соколов, А.Л. Крылова и Г.Г. Араке- 72 I.ыва I Нейрофизиология Клеточные основы обучения 7 Механизмы научения 73
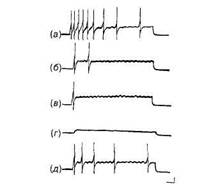
лов открыли феномен эндонейронального привыкания, в основе которого лежи г пластичность пеисмекерного механизма. Электрофизиологические характеристики пейсмекерных потенциалов, их роль в организации поведения, участие в процессах памяти изучались на полу-интактных препаратах и полностью изолированных нейронах. Особенностью пейсмекерной активности является ее генетическая заданность, а многообразие проявлений пейсмекерной активности нейрона дает возможность подойти к исследованию проблемы нейро-генетических механизмов памяти. Результаты, полученные в опытах А.Л Крыловой и Г.Г. Аракелова, сделали возможным предположение об эндонейрональной природе энграммы, формирующейся в процессе негативного обучения. В дальнейшем идеи об эндонейрональной природе пластичности пейсмекерных потенциалов были подтверждены в экспериментах на полностью изолированных нейронах виноградной улитки. На этом препарате была воспроизведена вся основная феноменология, открытая исследователями на полуинтактных препаратах и препарате изолированной нервной системы, в том числе и привыкание (рис. 6).
Рис 6 Привыкание изолированного нейрона Helix pomatia к повторяющимся электрическим раздражениям через внутриклеточный микроэлектрод Сила тока 1,5 нА, длительность импульса 10 с а—в — ответы на 1, 5 и 9 предъявление, г—ответ на применение деполяризационного тока силой 0,6 нА (экстрастимул), д — ответ на стимул прежней силы после предъявления экстрастимула (частичное растормаживание) Калибровка ЮмВ, 1 с Детальное изучение внутриклеточных механизмов привыкания и ассоциативного обучения предполагает использование нейронов, позволяющих не только вводить в них микроэлектроды, но и локально апплицировать биологически активные вещества. Для проверки гипотезы о внутриклеточном механизме обучения важно по- лучать изолированные нейроны. Всем этим условиям удовлетворяют гигантские нейроны моллюсков. В поведенческих опытах было показано, что у моллюсков можно выработать как привыкание, так и более сложные формы обучения. Однако, прежде чем описывать пластические изменения в идентифицированных нейронах моллюска, необходимо рассмотреть некоторые общие закономерности их развития. Селективность привыкания. Наличие двух пейсмекерных потенциалов, каждый из которых запускает свой независимый ПД фиксированной амплитуды и следовой гиперполяризации, позволяет изучить степень селективности развития привыкания. Повторение применения деполяризующего тока постепенно перестает вызывать реакцию одного пеисмекерного локуса, но продолжает увеличивать частоту другого пеисмекерного потенциала и связанного с ним ПД. Следует обратить внимание на то, что степень адаптации при непрерывном раздражении не коррелирует со скоростью развития привыкания. Сильная адаптация в одном локусе может сопровождаться слабым привыканием к току. Наоборот, локус со слабо выраженной адаптацией при многократном повторении раздражений может давать эффективное привыкание к внутриклеточному раздражению. Таким образом, локальность адаптации может не совпадать с локальными процессами, ответственными за привыкание. Развитие привыкания в одном из локусов может определяться: 1) выработкой компенсации сдвига мембранного потенциала с постстимульной гиперполяризацией; 2) выработкой локального изменения плотности тока в одном из пейсмекерных локусов; 3) локальной компенсацией действующего тока с торможением пейсмекерной активности, но без сдвига мембранного потенциала. Перерыв, введенный между стимулами, приводит к тому, что реакция восстанавливается и требуется повторение раздражений, чтобы опять получить привыкание. Нанесение более сильного стимула не только само вызывает реакцию, но и приводит к локальному восстановлению реакции на более слабый раздражитель, который ранее использовался в опыте для угашения реакций. Последнее обстоятельство имеет особое значение как воспроизведение эффекта «рас-тормаживания», характерного для поведенческого привыкания. Действительно, при развитии утомления, сенсорной адаптации или десенситизацни более сильный раздражитель лишь подавляет действие более слабого Для проверки селективности привыкания используется применение тока противоположного знака — изменение направления тока не только вызывает реакцию в локусе, подверга- 74 Нейрофизиология Клеточные основы обучения 7 Механизмы научения 75
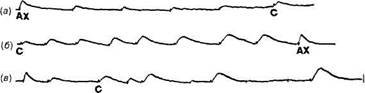
Многократное повторение гиперполяризующих импульсов тока приводит к тому, что величина реакции, заключающейся в подавлении пейсмекерной активности, постепенно уменьшается. Привыкание к гиперполяризующему току развивается преимущественно в одном из пейсмекерных локусов. Это также характеризует селективность в развитии привыкания. Нанесение более сильного раздражителя, однократная замена тока по знаку — все это вызывает растормаживание, как и в случае деполяризующего тока. Для изучения взаимодействия ПСП и пейсмекерных потенциалов можно имитировать ВПСП и ТПСП при помощи импульсов тока, поскольку в этом случае сопротивление мембраны остается постоянным. Регулярные импульсы тока, подаваемые через введенный в клетку микроэлектрод, вызывают сдвиг мембранного потенциала, регистрируемый вторым внутриклеточным микроэлектродом. Наиболее простой формой взаимодействия постсинаптических потенциалов (ПСП) с пейсмекерным потенциалом является случай, когда происходит линейная суммация пейсмекерного потенциала с вызванным сдвигом мембранного потенциала, имитирующего ПСП. В чистом виде такая суммация может наблюдаться в том случае, когда генерация ПД выключена. Функция постсинаптических структур в процессах обучения и памяти с наибольшей наглядностью проявлялась в опытах на полностью изолированных нейронах — ввиду особенностей данной биологической модели все пластические изменения связаны только с соматическим отделом нейрона и часть из них имеет эндонейрональную локализацию. Например, в опытах на изолированных клетках работа синапса моделируется при помощи микроионофоретически наносимого в хе-мочувствительные участки мембраны нейромедиатора. Повторное нанесение медиатора в один и тот же локус может приводить к развитию десенситизации (постепенной потере чувствительности мембраны к действующему нейромедиатору) или сенситизации (повышению чувствительности к нейромедиатору) (рис. 7, а). Удивительно, что хемо-чувствительные локусы мембраны одного и того же нейрона могут демонстрировать разнонаправленные изменения при повторных применениях медиаторов (рис. 7, б). Еще более удивительно, что такие изменения могут развиваться независимо одно от другого при одновременной (несинхронизированной) стимуляции сразу двух локусов (рис. 7, в).
АХ С Рис 7 Пластичность ответов на нейромедиаторы ацетилхолин (АХ) и серотонин (С), апплицируемые к изолированной соме идентифицированного нейрона ЛПа2 виноградной улитки а — развитие десенситизации при повторных предъявлениях АХ, б — сенси-тизация при повторных действиях С, в — развитие десенситизации и сенситизации при одновременном проведении стимуляции АХ и С Калибровка' 10 мВ, 1 с Многообразие следовых эффектов, обнаруживаемых в опытах на полностью изолированных нервных клетках, простота формирования ассоциативных связей между действующими нейрохимическими и электрическими воздействиями, поступающими на нейрон в опытах, моделирующих различные ситуации обучения, демонстрируют исключительно высокую степень пластичности, потенциально заложенную в каждую нервную клетку. По-видимому, в дальнейшем, вступая в нейронные связи, создаваемые мозгом по определенным критериям, нейроны реализуют тот пластический потенциал, который они изначально имели до формирования системы. Два возможных механизма постсинаптической пластичности. Опыты с ионофоретической аппликацией АХ на соматическую мембрану нейрона приводят к представлению о двух механизмах постсинаптической пластичности. Проявлением первого механизма является постсинаптическое изменение чувствительности рецепторов мембраны. Второй механизм проявляется в ослаблении действия вызванного медиатором синаптического ВПСП на пейсмекерную активность. Чрезвычайно интересным представляется тот факт, что изменение частоты генерации пейсмекерных ПД и смещение уровня МП при привыкании и фасилитации к микроаппликации медиатора могут протекать относительно независимо Это свидетельствует о том, что, по-видимому, в основе привыкания пейсмекерных ПД к постоянным сдвигам МП лежит особый механизм, отличный от десенситизации. Возможно, оно происходит за счет уменьшения влияний вызванного АХ ионного тока на пейсмекерный локус, как это имеет место при внутриклеточной инъекции ионов. Особый эффект аппликации АХ 76 Г пава I Нейрофизиология Клеточные основы обучения 7 Механизмы научения /7
Химическая чувствительность сомы нейрона к медиатору имеет существенное биологическое значение. Ученые считают, что кроме обычной синаптической передачи существует экстрасинаптическая реализация медиатора, не опосредованная специфической анатомической организацией. Очевидно, в этом случае передатчик, диффундируя в межклеточное пространство и действуя прямо на сому нейрона, снабженную адекватными хеморецепторами, тем самым модулирует нейронную активность. Исходя из этой гипотезы, нельзя исключить возможность прямого нейрогуморального влияния на пейсмекерный локус и в естественных условиях. Таким образом, реакции, полученные в этих опытах, являются не только аналогами синаптических ответов, но, возможно, отражают и естественную для них форму передачи возбуждения экстрасинаптическим путем.
_ h _
(д) Рис 8 Изменение амплитуды ПД при повторении деполяризационного стимула силой 0,35 мВ, длительностью 1 с (а — д) Стимул предъявляется с частотой 1 раз в 15 е — восстановление амплитуды ПД через 1 мин перерыва в предъявлении стимулов Калибровка 10 мВ 1с. Факты, свидетельствующие о селективном угасании реакций в разных участках соматической мембраны, доказывают, чтр нейрон привыкает к раздражителю не как единое целое, а как сложная система гетерогенных локусов, которые могут изменять свою реакцию на раздражитель независимо друг от друга. 7.3. Пластичность нейронов Классические условные рефлексы первым описал в начале XX в. И.П. Павлов. При классическом условном рефлексе исходно неэффективный раздражитель, называемый условным (точнее его следовало бы назвать раздражителем, который должен стать условным), повторно сочетается с высокоэффективным раздражителем, называемым безусловным. Вначале условный раздражитель вызывает лишь слабый ответ или не вызывает вообще никакого ответа; безусловный раздражитель провоцирует бурную реакцию без какого бы то ни было предварительного обучения. В результате выработки условного рефлекса (обучения) условный раздражитель приобретает способность вызывать либо более сильный, либо новый ответ. Для того, чтобы образовалась условная связь, произошло обучение, условный раздражитель должен коррелировать с безусловным, предшествуя ему на некоторый критический промежуток времени. Считается, что в результате животное обуч
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-01-09; просмотров: 208; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.102 (0.27 с.) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||





 ннем критериев начала реакции при ослаблении депрессии альфа-ритма.
ннем критериев начала реакции при ослаблении депрессии альфа-ритма.


 чтобы установить конфигурацию следа, возникшего в результате многократных нанесений стимула эталона.
чтобы установить конфигурацию следа, возникшего в результате многократных нанесений стимула эталона.



 кает, если же не совпадает, то ориентировочная реакция возникает, причем она тем интенсивнее, чем больше различий между новым стимулом и следом, оставленным в результате многократного нанесения предшествующих стимулов. Конфигурация следа, оставленного в нервной системе в результате повторения раздражителя, фиксирует все параметры сигнала. Можно сказать, что нервная система модификацией своих элементов строит нервную модель внешнего стимула.
кает, если же не совпадает, то ориентировочная реакция возникает, причем она тем интенсивнее, чем больше различий между новым стимулом и следом, оставленным в результате многократного нанесения предшествующих стимулов. Конфигурация следа, оставленного в нервной системе в результате повторения раздражителя, фиксирует все параметры сигнала. Можно сказать, что нервная система модификацией своих элементов строит нервную модель внешнего стимула.







 ющемся действию многократного раздражения, но и восстанавливает реакцию на ток того знака, который применяли в основном опыте по привыканию. Таким образом, смена направления тока также вызывает растормаживание.
ющемся действию многократного раздражения, но и восстанавливает реакцию на ток того знака, который применяли в основном опыте по привыканию. Таким образом, смена направления тока также вызывает растормаживание.
 состоит в ускорении или замедлении пейсмекерных ПД без сдвига МП нейрона. В связи со случаями подавления или усиления пейсме-керной активности без признаков изменения МП при прямом воздействии АХ на мембрану сомы, аналогичными действию «ускоряющих» или «замедляющих» синапсов, возникает предположение об особой чувствительности пейсмекерного локуса к медиатору.
состоит в ускорении или замедлении пейсмекерных ПД без сдвига МП нейрона. В связи со случаями подавления или усиления пейсме-керной активности без признаков изменения МП при прямом воздействии АХ на мембрану сомы, аналогичными действию «ускоряющих» или «замедляющих» синапсов, возникает предположение об особой чувствительности пейсмекерного локуса к медиатору.
 Важным фактом, заставляющим связывать пластические изменения с деятельностью постсинаптических структур, являются изменения, развивающиеся при повторных предъявлениях внутриклеточных импульсов тока. Из наших экспериментов следует, что пластические изменения при микроаппликации АХ могут быть аналогичны полученным при внутриклеточной стимуляции и развиваться при постоянной чувствительности хеморецепторов мембраны. Механизмы привыкания при инъекции ионов и аппликации АХ оказываются связанными с пейсмекерным потенциалом. Привыкание и фасилитация в звене пейсмекерного механизма могут развиваться и без сдвига МП. Эти формы привыкания и фасилитации также отличаются от явлений сенситизации и десенситизации рецепторов, при которых наблюдается уменьшение и полное исчезновение вызванных медиатором сдвигов МП (рис. 8).
Важным фактом, заставляющим связывать пластические изменения с деятельностью постсинаптических структур, являются изменения, развивающиеся при повторных предъявлениях внутриклеточных импульсов тока. Из наших экспериментов следует, что пластические изменения при микроаппликации АХ могут быть аналогичны полученным при внутриклеточной стимуляции и развиваться при постоянной чувствительности хеморецепторов мембраны. Механизмы привыкания при инъекции ионов и аппликации АХ оказываются связанными с пейсмекерным потенциалом. Привыкание и фасилитация в звене пейсмекерного механизма могут развиваться и без сдвига МП. Эти формы привыкания и фасилитации также отличаются от явлений сенситизации и десенситизации рецепторов, при которых наблюдается уменьшение и полное исчезновение вызванных медиатором сдвигов МП (рис. 8).


