
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Основные физиологические категорииСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Живые организмы представляют собой открытые системы, т.е. неразрывно связанные с внешней средой. К числу закономерностей, совокупность которых характеризует жизнь, относятся: ■ самообновление; ■ саморегуляция (авторегуляция); ■ самовоспроизведение. Перечисленные закономерности обусловливают основные атрибуты жизни (основные свойства живого организма): обмен веществ, раздражимость (возбудимость), подвижность, размножение, наследственность, поддержание гомеостаза, приспособляемость (адаптивность). Процесс обмена веществ у всех живых организмов, начиная от самых примитивных и кончая самым сложным - человеческим организмом, является одной из главных основ жизни. Он осуществляется сложнейшими химическими соединениями, которые называются белками. Ф. Энгельс писал, что «жизнь есть способ существования белковых тел, существенным моментом которого является обмен веществ с окружающей их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь, что приводит к разложению белка». Происходящие в организме физиологические процессы сводятся, в конечном счете, к обеспечению обмена веществ и его регуляции в соответствии
с потребностями организма, которые определяются условиями его жизни. Для отправления этих функций существуют многочисленные специальные органы, находящиеся в непрерывном и очень сложном взаимодействии. По словам английского ученого Д. Уолда «самая сложная машина из всех, какие когда-либо создавал человек – ну, скажем, электронный мозг – не более чем детская игрушка по сравнению с самым простым из живых существ». Человек же, как известно, это самое сложное из живых существ. Каждая из ста триллионов клеток организма человека отличается чрезвы-чайно сложной структурой, способностью к самоорганизации и многосто-роннему взаимодействию с другими клетками. Клетка же представляет со-бой лишь одну из сравнительно элементарных подсистем в сложной ие-рархии систем, формирующих живой организм. Нормальное функционирование организма и каждого отдельного элемента системы (в т.ч. и каждой клетки) возможно благодаря непрерыв-ному обмену информацией между элементами (и между клетками). 2. Гомеостаз Внутренняя среда организма, в которой живут все его клетки, – это кровь, лимфа, межтканевая жидкость. Ее характеризует относительное по-стоянство – гомеостаз различных показателей, т.к. любые ее изменения приводят к нарушению функций клеток и тканей организма, особенно вы-сокоспециализированных клеток центральной нервной системы. К таким постоянным показателям гомеостаза относятся температура внутренних органов тела, сохраняемая в пределах 36 – 37 ºС, кислотно-основное рав-новесие крови, характеризуемое величиной рН = 7,4 – 7,35, осмотическое давление крови (7,6 – 7,8 атм), концентрация гемоглобина в крови – 120 – 140 г/ л и др. Гомеостаз представляет собой не статическое явление, а динамиче-ское равновесие. Способность сохранять гомеостаз в условиях постоянно-го обмена веществ и значительных колебаний факторов внешней среды обеспечивается комплексом регуляторных реакций организма. Эти регуля-торные процессы поддержания динамического равновесия получили на-звание гомеокинеза. Степень сдвига показателей гомеостаза при существенных колеба-ниях условий внешней среды или при тяжелой работе у большинства лю-дей очень невелика. Например, длительное изменение рН крови всего на 0,1 – 0,2 может привести к смертельному исходу. Однако в общей популя-ции имеются отдельные индивиды, обладающие способностью переносить гораздо большие сдвиги показателей внутренней среды. У высококвали-фицированных спортсменов-бегунов в результате большого поступления молочной кислоты из скелетных мышц в кровь во время бега на средние и длинные дистанции рН крови может снижаться до величин 7,0 и даже 6,9. Лишь несколько человек в мире оказались способными подняться на высо-ту порядка 8 800 м над уровнем моря (на вершину Эвереста) без кислород-ного прибора, т.е. существовать и двигаться в условиях крайнего недостат-ка кислорода в воздухе и, соответственно, в тканях организма. Эта способ-ность определяется врожденными особенностями человека – так называе-мой его генетической нормой реакции, которая даже для достаточно по-стоянных функциональных показателей организма имеет широкие инди-видуальные различия. 3. Нервные и гуморальные механизмы регуляции функций Обмен информацией в живых организмах происходит посредством прямого (контактного) взаимодействия между клетками, в результате транспорта веществ с тканевой жидкостью, лимфой и кровью (гуморальная связь – от лат. гумор – жидкость), а также при передаче от клетки к клетке биоэлектрических потенциалов, что представляет самый быстрый способ передачи информации в организме. У многоклеточных организмов разви-лась специальная система, обеспечивающая восприятие, передачу, хране-ние, переработку и воспроизведение информации, закодированной в элек-трических сигналах. Это – нервная система, достигшая у человека наи-высшего развития. Таким образом, регуляция различных функций у высокооргани - зованных животных и человека осуществляется, в основном, двумя путями: гуморальным ( через кровь, лимфу и тканевую жидкость ) и нервным. Возможности гуморальной регуляции ограничены тем, что она дей-ствует сравнительно медленно и не может обеспечить срочных ответов ор-ганизма (быстрых движений, мгновенной реакции на экстренные раздра-жители). Кроме того, гуморальным путем происходит широкое вовлечение различных органов и тканей в реакцию. В отличие от этого, с помощью нервной системы возможно быстрое и точное управление различными отделами целостного организма, доставка сообщений точному адресату. Оба эти механизма тесно связаны, однако ведущую роль в регуля - ции функций играет нервная система. Нервная система человека функционирует как сложное кибернетиче-ское устройство, равного которому нет ни в природе, ни в технике. Непре- рывно получая колоссальный поток информации, поступающий от внут-ренних органов, сосудов, аппарата движения и из внешней среды – через органы чувств, – она регулирует и согласовывает деятельность всех орга-нов в интересах всего организма в целом. Нервная система регулирует весь режим его внутренней жизни в соответствии с условиями внешней среды. В регуляции функционального состояния органов и тканей прини-мают участие особые вещества – нейропептиды, выделяемые гипофизом и нейронами спинного и головного мозга. Известно около сотни подобных веществ, которые, не вызывая сами возбуждения клеток, могут заметно изменять их функциональное состояние. Они влияют на сон, процессы обучения и памяти, на мышечный тонус, вызывают обездвижение или об-ширные судороги мышц, обладают обезболивающим и наркотическим эф-фектом. Концентрация нейропептидов в крови у спортсменов может пре-вышать средний уровень у нетренированных лиц в 6 – 8 раз, повышая эф-фективность соревновательной деятельности. Чрезмерные тренировочные занятия вызывают истощение нейропептидов и срыв адаптации спортсме-на к физическим нагрузкам.
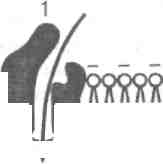
Лекция 3 ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ. 1. Биоэлектрическая активность живых клеток. Основные функциональные характеристики возбудимых тканей. 2. Мембранный потенциал. Природа потенциала покоя. 3. Калий-натриевый насос мембраны. 4. Потенциал действия. Ионный механизм возникновения потенциала действия. 5. Проведение возбуждения. 1. Биоэлектрическая активность живых клеток. Основные функциональные характеристики возбудимых тканей Чтобы понять природу биоэлектрических явлений, т.е. сигналов, при помощи которых нервная система осуществляет передачу информации, необходимо прежде всего рассмотреть некоторые стороны общей физиологии так называемых возбудимых тканей, к которым относятся нервная, мышечная и железистая ткань. Все живые клетки обладают раздражимостью, т.е. способностью под влиянием определенных факторов внешней или внутренней среды, так называемых раздражителей, переходить из состояния физиологического покоя в состояние активности. Однако термин возбудимые клетки применяют лишь по отношению к нервным, мышечным и секреторным клеткам, способным в ответ на действие раздражителя генерировать электрические потенциалы. Рассмотрим основные функциональные характеристики возбудимых тканей: возбудимость, проводимость и лабильность. Возбудимость – свойство возбудимых тканей отвечать на раздражение специфическим процессом возбуждения. Этот процесс включает электрические, ионные, химические и тепловые изменения, а также специфические проявления: в нервных клетках – импульсы возбуждения, в мышечных – сокращение или напряжение, в железистых – выделение определенных веществ (секретов). Для нервной и мышечной ткани характерна также способность передавать активное состояние соседним участкам – т.е. проводимость (способность проводить возбуждение). Возбудимые ткани характеризуются двумя основными нервными процессами – возбуждением и торможением. Торможение – это активная задержка процесса возбуждения. Взаимодействие этих двух процессов обеспечивает координацию нервной деятельности в целостном организме. Различают местное (локальное) возбуждение и распространяющееся. Местное возбуждение представляет незначительные изменения в поверхностной мембране клеток, а распространяющееся – связано с передачей импульса возбуждения вдоль нервной или мышечной ткани. Для измерения возбудимости пользуются определением порога, т.е. минимальной величины раздражения, при которой возникает распространяющееся возбуждение. Величина порога зависит от функционального состояния ткани и от особенностей раздражителя, которым может быть любое изменение внешней среды (электрическое, тепловое, механическое и пр.). Чем выше порог, тем ниже возбудимость, и наоборот. Возбудимость может повышаться в процессе выполнения физических упражнений (например, под влиянием разминки, в ходе врабатывания) и снижаться при утомлении, развитии перетренированности. Лабильность – скорость протекания процесса возбуждения в нервной и мышечной тканях (лат. лабилис – подвижный). Понятие лабильности, или функциональной подвижности, было выдвинуто Н. Е. Введенским в 1892 г. В качестве одной из мер лабильности Введенский предложил максимальное количество волн возбуждения (электрических потенциалов действия), которое может воспроизводиться тканью в 1 с в соответствии с ритмом раздражения. Лабильность характеризует скоростные свойства. ткани. Она может повышаться под влиянием раздражений, тренировки, особенно у спортсменов при развитии качества быстроты. Первые данные о существовании биоэлектрических явлений («животное электричество») были получены в конце 18 в. при изучении природы электрического разряда, наносимого некоторыми рыбами при защите и нападении. Итальянский ученый Луиджи Гальвани установил наличие электрических потенциалов в нервной и мышечной тканях. Однако первые прямые измерения потенциалов в живых тканях стали возможны только после изобретения гальванометров. Систематическое исследование потенциалов в мышцах и нервах в состоянии покоя и возбуждения было начато в 1848 г. Э. Дюбуа Рейманом. Дальнейшие успехи в изучении биоэлектрических явлений были тесно связаны с усовершенствованием техники регистрации быстрых колебаний электрического потенциала (струнные, катодные, шлейфные осциллографы) и методов их отведения от одиночных возбудимых клеток. Качественно новый этап в изучении электрических явлений в живых тканях – 40 – 50 гг. 20 в. С помощью внутриклеточных микроэлектродов удалось произвести прямую регистрацию электрических потенциалов клеточных мембран. В основе колебаний электрических потенциалов лежат обратимые изменения проницаемости клеточной мембраны для определенных ионов. В свою очередь изменение проницаемости является следствием открывания и закрывания существующих в клеточной мембране ионных каналов под влиянием действующего раздражителя. Энергия, используемая при генерации электрических потенциалов, запасена в покоящейся клетке в виде градиентов концентраций ионов Na+, Cа 2+, К+, Сl- по обе стороны поверхностной мембраны. Указанные градиенты создаются и поддерживаются работой специализированных молекулярных устройств, так называемых мембранных ионных насосов. Последние используют для своей работы энергию обмена веществ, выделяющуюся при ферментативном расщеплении универсального клеточного донатора энергии – аденозинтрифосфорной кислоты (АТФ). Изучение электрических потенциалов, сопровождающих процессы возбуждения и торможения в живых тканях, имеет важное значение как для понимания природы этих процессов, так и для выявления характера нарушений деятельности возбудимых клеток при различных видах патологии. 2. Мембранный потенциал. Природа потенциала покоя Термином «мембранный потенциал» (потенциал покоя) принято называть трансмембранную разность потенциалов, существующую между цитоплазмой и окружающим клетку наружным раствором (между внешней и внутренней поверхностью мембраны). Когда клетка (волокно) находится в состоянии физиологического покоя, ее внутренний потенциал отрицателен по отношению к наружному, условно принимаемому за нуль. У различных клеток мембранный потенциал варьирует от -50 до -90 мВ. Чтобы измерить потенциал покоя и проследить его изменения, вызываемые тем или иным воздействием на клетку, применяют технику внутриклеточных микроэлектродов. Как только микроэлектрод прокалывает поверхностную мембрану клетки, луч осциллографа сразу же отклоняется от своего исходного (нулевого) положения, обнаруживая тем самым существование разности потенциалов между поверхностью и содержимым клетки. Дальнейшее продвижение микроэлектрода внутри протоплазмы на положении луча осцилло-рафа не сказывается. Это свидетельствует о том, что потенциал действительно локализуется на клеточной мембране. Cуществует множество факторов, меняющих потенциал покоя клеток: приложение электрического тока, изменение ионного состава среды, воздействие токсинов, нарушение кислородного снабжения ткани и т.д. Во всех тех случаях, когда внутренний потенциал уменьшается (становится менее отрицательным), говорят о деполяризации мембраны, противоположный сдвиг потенциала (увеличение отрицательного заряда внутренней поверхности клеточной мембраны) называют гиперполяризацией. Природа потенциала покоя. Еще в 1896 г. В. Ю. Чаговец высказал гипотезу об ионном механизме электрических потенциалов в живых клетках. В 1902 г. Ю. Бернштейном была развита мембранно-ионная теория, которую модифицировали и экспериментально обосновали английские ученые А. Ходжкин, А. Хаксли и Б. Кац (1949 – 1952). Согласно указанной теории наличие электрических потенциалов в живых клетках обусловлено неравенством концентрации ионов Na+, К +, Ca2+ и Cl- внутри и вне клетки и различной проницаемостью для них поверхностной мембраны. Известно, что содержимое нервного волокна богато K+ и органическими анионами (практически не проникающими через мембрану) и бедно Na+ и Cl-. Концентрация К+ в цитоплазме нервных и мышечных клеток в 40 – 50 раз выше, чем в наружном растворе, и если бы мембрана в покое была проницаема только для этих ионов, то потенциал покоя соответствовал бы равновесному калиевому потенциалу Ек = -97,5 мВ, рассчитанному по формуле Нернста. Чтобы понять, каким образом возникает этот потенциал, рассмотрим следующий модельный опыт (рис. 1). Представим сосуд, разделенный искусственной полупроницаемой мембраной. Стенки пор этой мембраны заряжены электроотрицательно, поэтому они пропускают только катионы и непроницаемы для анионов. В обе половины сосуда налит солевой раствор, содержащий ионы К+, однако их концентрация в правой части сосуда выше, чем в левой. Вследствие этого концентрационного градиента ионы.
К+ начинают диффундировать из правой половины сосуда в левую, принося туда свой положительный заряд. Это приводит к тому, что непроникающие анионы начинают скапливаться у мембраны в правой половине
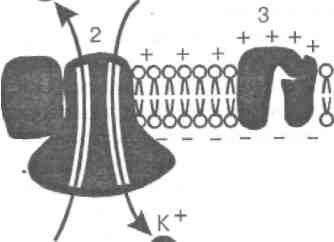
Своим отрицательным зарядом они электростатически будут удерживать К+ у поверхности мембраны в левой половине сосуда. В результате мембрана поляризуется, и между двумя ее поверхностями создается разность потенциалов, со-ответствующая равновесному калиевому потенциалу (Ек). Предположение о том, что в состоянии покоя мембрана нервных и мышечных волокон избирательно проницаема для К+, и что именно их диффузия создает потенциал покоя, было подтверждено А. Ходжкиным с сотрудниками в 1962 г. в опытах на изолированных гигантских аксонах кальмара. Из волокна диаметром около 1 мм осторожно выдавливали цитоплазму и спавшуюся оболочку заполняли искусственным солевым раствором. Когда концентрация К+ в растворе была близка к внутриклеточной, между внутренней и наружной сторонами мембраны устанавливалась разность потенциалов, близкая к значению нормального потенциала покоя (-50 - -70 мВ), и волокно проводило импульсы. Такие опыты показали, что градиент концентрации К+ действительно является основным фактором, определяющим величину потенциала покоя нервного волокна. Однако покоящаяся мембрана проницаема не только для К+, но (правда, в значительно меньшей степени) и для Na+. Диффузия этих положительно заряженных ионов внутрь клетки уменьшает абсолютную величину внутреннего отрицательного потенциала клетки, создаваемого диффузией К+. Поэтому потенциал покоя волокон (-50 - -70 мВ) менее отрицателен, чем рассчитанный по формуле Нернста. Ионы Cl- в нервных волокнах не играют существенной роли в образовании потенциала покоя, т.к. проницаемость для них покоящейся мембраны относительно мала. В отличие от этого в скелетных мышечных волокнах проницаемость мембраны для ионов хлора сравнима с калиевой, и поэтому диффузия Cl- внутрь клетки увеличивает значение потенциала покоя. Таким образом, мембрана в состоянии покоя поляризована, т. е. имеется определенная разность потенциалов по обе стороны мембраны, называемая потенциалом покоя. Она равна для нейрона примерно -70 мВ, для мышечного волокна – -90 мВ. 3. Калий - натриевый мембраны Несмотря на то что потоки К+ и Na+ через мембрану в покое малы, разность концентраций этих ионов внутри клетки и вне ее должна была бы в конечном итоге выровняться, если бы в клеточной мембране не существовало особого молекулярного устройства – калий-натриевого насоса, которое обеспечивает выведение (выкачивание) из цитоплазмы проникающих в нее Na+ и введение (нагнетание) в цитоплазму К+ (рис. 2). К" Na"
кдаш Бшжч рш
+ + + т + + + Na
Натриевый насос перемещает К+ и Na+ против их концентрационных градиентов, т.е. совершает определенную работу. Непосредственным источником энергии для этой работы является богатое энергией (макроэргическое) соединение – аденозинтрифосфат (АТФ), являющееся универсальным источ-ником энергии живых клеток. Расщепление АТФ производится ферментом АТФ-азой, локализованной в поверхностной мембране клетки.
Рис 2 Мембрана возбудимых клеток в покое (А) и при возбуждении (Б): а - двойной слой липидов, б - белки мембраны. На А: 1 - ионный калиевый канал, 2 - натрий-калиевый насос, 3 - закрытый в покое натриевый канал. На Б: 1 - открытый при возбуждении натриевый канал, 2 - вхождение ионов натрия в клетку и смена зарядов на наружной и внутренней стороне мембраны Роль АТФ в механизме активного транспорта Na+ и К+ прямо доказана в опытах на гигантских нервных волокнах кальмара. Было установлено, что путем введения внутрь волокна АТФ можно временно восстановить работу калий-натриевого насоса, нарушенную ингибитором дыхательных ферментов цианидом. Первоначально полагали, что натриевый насос электронейтрален, т.е. число обмениваемых ионов Na+ и К+ равно. В дальнейшем выяснилось, что на каждые три иона натрия, выводимые из клетки, в клетку поступает только два иона калия. Это означает, что калий-натриевый насос электрогенен: он создает на мембране разность потенциалов, суммирующуюся с потенциалом покоя. Таким образом, в формировании потенциала покоя натриевый насос играет двоякую роль: ■ создает и поддерживает трансмембранный градиент концентраций Na+ и К+; ■ генерирует разность потенциалов, суммирующуюся с потенциалом, создаваемым диффузией К+ по концентрационному градиенту.
|
||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-06; просмотров: 619; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.118.1.63 (0.01 с.) |

 сосуда.
сосуда.
 .
.


