Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Потенциал действия. Ионный механизм возникновения потенциала действияСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Потенциалом действия (ПД) называют быстрое колебание мембранного потенциала, возникающее при возбуждении нервных, мышечных и некоторых других клеток. В его основе лежат изменения ионной проницаемости мембраны. Амплитуда ПД мало зависит от силы вызывающего его раздражителя, важно лишь, чтобы эта сила была не меньше некоторой критической величины, которая называется порогом раздражения. Возникнув в месте раздражения, ПД распространяется вдоль нервного или мышечного волокна, не изменяя своей амплитуды. В естественных условиях ПД генерируются в нервных волокнах при раздражении рецепторов или возбуждении нервных клеток. Распространение ПД по нервным волокнам обеспечивает передачу информации в нервной системе. Достигнув нервных окончаний, ПД вызывают секрецию химических веществ (медиаторов), обеспечивающих передачу сигнала на мышечные или нервные клетки. В мышечных клетках ПД инициируют цепь процессов, вызывающих сократительный акт. Ионы, проникающие в цитоплазму во время генерации ПД, оказывают регулирующее влияние на метаболизм клетки и, в частности, на процессы синтеза белков, составляющих ионные каналы и ионные насосы.
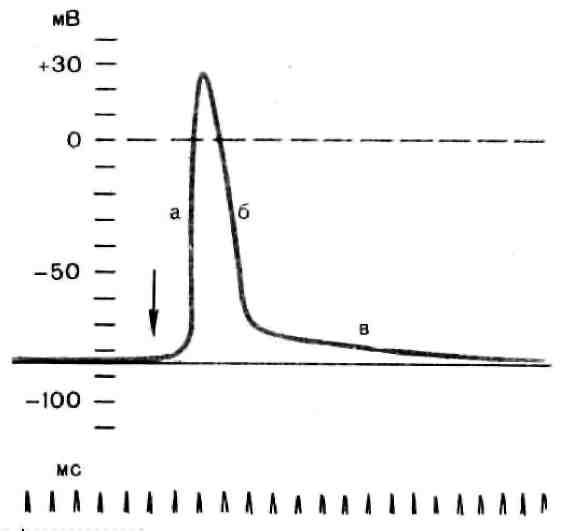
Рис. 3. Потенциал действия скелетного мышечного волокна, зарегистрированный с помощью внутриклеточного микроэлектрода: а – фаза деполяризации, б – фаза реполяризации, в – фаза следовой деполяризации (отрицательный следовой потенциал). Момент нанесения раздражения показан стрелкой. Установлено, что во время восходящей фазы (фазы деполяризации) происходит не просто исчезновение потенциала покоя (как это первоначально предполагали), а возникает разность потенциалов обратного знака: внутреннее содержимое клетки становится заряженным положительно по отношению к наружной среде, иными словами, происходит реверсия мембранного потенциала. Во время нисходящей фазы (фазы реполяризации) мембранный потенциал возвращается к своему исходному значению. Если рассмотреть пример записи ПД в скелетном мышечном волокне лягушки (см. рис. 3), то видно, что в момент достижения пика мембранный потенциал составляет +30 – +40 мВ. Длительность пика ПД у различных нервных и мышечных волокон варьирует от 0,5 до 3 мс, причем фаза реполяризации продолжительнее фазы деполяризации. Изменения мембранного потенциала, следующие за пиком потенциала действия, называют следовыми потенциалами. Различают два вида следовых потенциалов – следовую деполяризацию и следовую гиперполяризацию. Ионный механизм возникновения ПД. Как отмечалось, в состоянии покоя проницаемость мембраны для калия превышает ее проницаемость для натрия. Вследствие этого поток К+ из цитоплазмы во внешний раствор превышает противоположно направленный поток Na+. Поэтому наружная сторона мембраны в покое имеет положительный потенциал по отношению к внутренней. При действии на клетку раздражителя проницаемость мембраны для Na+ резко повышается и становится примерно в 20 раз больше проницаемости для K+. Поэтому поток Na+ из внешнего раствора в цитоплазму начинает превышать направленный наружу калиевый ток. Это приводит к изменению знака (реверсии) мембранного потенциала: внутренняя сторона мембраны в месте возбуждения становится заряженной положительно по отношению к ее наружной поверхности. Указанное изменение мембранного потенциала соответствует восходящей фазе ПД (фазе деполяризации). Повышение проницаемости мембраны для Na+ продолжается лишь очень короткое время. Вслед за этим проницаемость мембраны для Na+ вновь понижается, а для K+ возрастает. Процесс, ведущий к понижению ранее увеличенной натриевой проницаемости мембраны, назван натриевой инактивацией. В результате инактивации поток Na+ внутрь цитоплазмы резко ослабляется. Увеличение же калиевой проницаемости вызывает усиление потока K+ из цитоплазмы во внешний раствор. В итоге этих двух процессов и происходит реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к наружной стороне мембраны. Этому изменению потенциала соответствует нисходящая фаза ПД (фаза реполяризации). Опыты на гигантских нервных волокнах кальмара позволили получить подтверждение правильности натриевой теории возникновения ПД. ПД возникает при деполяризации поверхностной мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению ионов Na внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные изменения на мембране ( локальный ответ ). При увеличении силы раздражения, когда достигнут порог возбудимости, изменения мембранного потенциала достигают критического уровня деполяризации (КУД). Например, величина потенциала покоя равна -70 мВ, КУД = -50 мВ. Чтобы вызвать возбуждение, надо деполяризовать мембрану до -50 мВ, т.е. на -20 мВ снизить ее исходный потенциал покоя. Только при достижении КУД наблюдается резкое изменение мембранного потенциала, которое регистрируется в виде ПД. Таким образом, основное условие возникновения потенциала действия – это снижение мембранного потенциала до критического уровня деполяризации. В основе рассмотренных изменений ионной проницаемости мембраны при генерации ПД лежат процессы открывания и закрывания специализированных ионных каналов в мембране, обладающих двумя важнейшими свойствами: ■ избирательностью (селективностью) по отношению к определенным ионам; ■ электровозбудимостью, т.е. способностью открываться и закрываться в ответ на изменения мембранного потенциала. Так же как ионные насосы, ионные каналы образованы макромолекулами белков, пронизывающими липидный бислой мембраны. Активный и пассивный ионный транспорт. В процессе восстановления после ПД работа калий-натриевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, благодаря чему нарушенное при возбуждении неравенство концентраций Na+ и K+ по обе стороны мембраны восстанавливается. На работу этого механизма тратится около 70 % необходимой клетке энергии. Таким образом, в живой клетке существует две системы движения ионов через мембрану. Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии (пассивный ионный транспорт). Он ответствен за возникновение потенциала покоя и ПД и ведет в конечном итоге к выравниванию концентрации ионов по обе стороны клеточной мембраны. Второй осуществляется против концентрационного градиента. Он состоит в «выкачивании» ионов натрия из цитоплазмы и «нагнетании» ионов калия внутрь клетки. Этот тип ионного транспорта возможен лишь при условии затраты энергии обмена веществ. Его называют активным ионным транспортом. Он ответствен за поддержание постоянства разности концентраций ионов между цитоплазмой и омывающей клетку жидкостью. Активный транспорт - результат работы натриевого насоса, благодаря которому восстанавливается исходная разность ионных концентраций, нарушающаяся при каждой вспышке возбуждения. Проведение возбуждения Нервный импульс (потенциал действия) обладает способностью распространяться вдоль по нервным и мышечным волокнам. В нервном волокне потенциал действия является очень сильным раздражителем для соседних участков волокна. Амплитуда потенциала действия обычно в 5 – 6 раз превышает пороговую величину деполяризации. Это обеспечивает высокую скорость и надежность проведения. Между зоной возбуждения (имеющей на поверхности волокна отрицательный заряд и на внутренней стороне мембраны – положительный) и соседним невозбужденным участком мембраны нервного волокна (с обратным соотношением зарядов) возникают электрические токи – так называемые местные токи. В результате развивается деполяризация соседнего участка, увеличение его ионной проницаемости и появление потенциала действия. В исходной же зоне возбуждения восстанавливается потенциал покоя. Затем возбуждением охватывается следующий участок мембраны и т.д. Таким образом, с помощью местных токов происходит распространение возбуждения на соседние участки нервного волокна, т.е. проведение нервного импульса. По мере проведения амплитуда потенциала действия не уменьшается, т.е. возбуждение не затухает даже при большой длине нерва. В процессе эволюции с переходом от безмякотных нервных волокон к мякотным (покрытым миелиновой оболочкой) произошло существенное повышение скорости проведения нервного импульса. Для безмякотных волокон характерно непрерывное проведение возбуждения, которое охватывает последовательно каждый соседний участок нерва. Мякотные же нервы почти полностью покрыты изолирующей миелиновой оболочкой. Ионные токи в них могут проходить только в оголенных участках мембраны -перехватах Ранвье, лишенных этой оболочки. При проведении нервного импульса потенциал действия перескакивает от одного перехвата к другому и может охватывать даже несколько перехватов. Такое проведение поучило название сальтоторного (лат. сальто – прыжок). При этом повышается не только скорость, но и экономичность проведения. Возбуждение захватывает не всю поверхность мембраны волокна, а лишь небольшую ее часть. Следовательно, меньше энергии тратится на активный транспорт ионов через мембрану при возбуждении и в процессе восстановления. Скорость проведения в разных волокнах различна. Более толстые нервные волокна проводят возбуждение с большей скоростью: у них расстояния между перехватами Ранвье больше и длиннее скачки. Наибольшую скорость проведения имеют двигательные и проприоцептивные афферентные нервные волокна - до 100 м/с. В тонких симпатических нерв ных волокнах (особенно в немиелинизированных волокнах) скорость проведения мала - порядка 0,5 - 15 м/с. Во время развития потенциала действия мембрана полностью теряет возбудимость. Это состояние называют полной невозбудимостью, или абсолютной рефрактерностью. За ним следует относительная рефрактерность, когда потенциал действия может возникать лишь при очень сильном раздражении. Постепенно возбудимость восстанавливается до исходного уровня. Законы проведения возбуждения в нервах: 1. Проведение импульсов возможно лишь при условии анатомической и физиологической целостности волокна. 2. Двустороннее проведение: при раздражении нервного волокна возбуждение распространяется по нему и в центробежном, и в центростремительном направлениях. 3. Изолированное проведение: в периферическом нерве импульсы распространяются по каждому волокну изолированно, т.е. не переходя с одного волокна на другое и оказывая действие только на те клетки, с которыми контактируют окончания данного нервного волокна.
13. Дайте определение гомеостаза. 14. Назовите основные пути регуляции различных функций у высокоорганизованных животных и человека. 15. Кем и когда было открыто «животное электричество»? 16. Какие ткани относятся к возбудимым? Почему они так называются? 17. Назовите основные функциональные характеристики возбудимых тканей. 18. Что называют порогом возбудимости? 19. От каких факторов зависит величина порога? 20. Что такое лабильность? Кем было выдвинуто понятие лабильности, какие свойства возбудимых тканей оно характеризует? 21. Что называют мембранным потенциалом (потенциалом покоя)? 22. Чем обусловлено наличие электрических потенциалов в живых клетках? 23. В каких случаях говорят о деполяризации (или гиперполяризации) клеточной мембраны? 24. Какую роль в формировании потенциала покоя играет калий-натриевый насос мембраны? 25. Что называют потенциалом действия? Какова его роль в нервной системе? 26. Что лежит в основе возникновения потенциала действия? 27. Охарактеризуйте фазы потенциала действия. 28. Что называют реверсией мембранного потенциала? 29. Опишите ионный механизм возникновения потенциала действия. 30. Что понимают под натриевой инактивацией? 31. Что такое критический уровень деполяризации? 32. Какими свойствами обладают ионные каналы клеточной мембраны? 33. Охарактеризуйте два типа ионного транспорта в клетке:
■ пассивный; ■ активный.
Модуль 1 ОБЩАЯ ФИЗИОЛОГИЯ ЦНС
|
|||||
|
|
Последнее изменение этой страницы: 2016-08-06; просмотров: 6188; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.117.151.198 (0.008 с.) |