Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Физиологический механизм восприятия звукаСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Восприятие звука основано на двух процессах, происходящих в улитке: ■ разделении звуков различной частоты по месту их наибольшего воздействия на основную мембрану улитки; ■ преобразовании рецепторными клетками механических колебаний в нервное возбуждение. Звуковые колебания, поступающие во внутреннее ухо через овальное окно, передаются перилимфе, а колебания этой жидкости приводят к смещениям основной мембраны, на которой расположены рецепторные волосковые клетки: внутренние и наружные, отделенные друг от друга кортиевыми дугами. Волоски рецепторных клеток омываются эндолимфой и контактируют с покровной мембраной, которая по всему ходу перепончатого канала расположена над волосковыми клетками. При действии звуков основная мембрана начинает колебаться, волоски рецепторных клеток касаются покровной мембраны и механически раздражаются. В результате в них возникает процесс возбуждения, который по афферентным волокнам направляется к нейронам спирального узла улитки и далее в ЦНС. От высоты звука зависит высота столба колеблющейся жидкости и, соответственно, место наибольшего смещения основной мембраны: звуки высокой частоты дают наибольший эффект на начале основной мембраны, а низких частот – доходят до вершины улитки. Таким образом, при различных по частоте звуках возбуждаются разные волосковые клетки и разные волокна. Увеличение силы звука приводит к увеличению числа возбужденных волосковых клеток и нервных волокон, что позволяет различать интенсивность звуковых колебаний. Различают костную и воздушную проводимость звука. В обычных условиях у человека преобладает воздушная проводимость – проведение звуковых колебаний через наружное и среднее ухо к рецепторам внутреннего уха. В случае костной проводимости звуковые колебания передаются через кости черепа непосредственно улитке (например, при нырянии, подводном плавании). Человек обычно воспринимает звуки с частотой от 15 до 20 000 Гц. У детей верхний предел достигает 22 000 Гц, с возрастом он понижается. Наиболее высокая чувствительность обнаружена в области частот от 1 000 до 3 000 Гц. Эта область соответствует наиболее часто встречающимся частотам человеческой речи и музыки.
4. Значение и общий план организации вестибулярной сенсорной системы Вестибулярная сенсорная система служит для анализа положения и движения тела в пространстве. Это одна из древнейших сенсорных систем, развившаяся в условиях действия силы тяжести на Земле. Наряду со зрительной сенсорной системой и кинестетическим анализатором она играет ведущую роль в пространственной ориентировке человека. Импульсы от вестибулорецепторов используются в организме для поддержания равновесия тела, для регуляции и сохранения позы, для пространственной организации движений человека. При равномерном движении или в условиях покоя рецепторы вестибулярной сенсорной системы не возбуждаются. Вестибулярная сенсорная система состоит из следующих отделов: 1. периферического, который включает два образования, содержащих механорецепторы вестибулярной системы, – преддверие (мешочек и маточка) и полукружные каналы; 2. проводникового, который начинается от рецепторов волокнами биполярной клетки ( первого нейрона ) вестибулярного узла, расположенного в височной кости, аксоны этих нейронов образуют вестибулярный нерв и вместе со слуховым нервом в составе 8-ой пары черепномозговых нервов входят в продолговатый мозг; в вестибулярных ядрах продолговатого мозга находятся вторые 3. нейроны, импульсы от которых поступают к третьим нейронам – в таламусе. Сигналы от вестибулярных ядер направляются не только к таламусу (это не единственный путь), они направляются во многие отделы ЦНС: спинной мозг, мозжечок, ретикулярную формацию и вегетативные ганглии. 3. коркового, представленного четвертыми нейронами, часть которых расположена в первичном поле вестибулярной системы в височной области коры, а другая – в непосредственной близости к пирамидным нейронам моторной области коры и в постцентральной извилине. Точная локализация вестибулярной зоны коры человека в настоящее время окончательно не выяснена. 5. Функционирование вестибулярного аппарата Итак, периферическим отделом вестибулярной сенсорной системы является вестибулярный аппарат, находящийся во внутреннем ухе в лабиринте пирамиды височной кости. Он состоит из преддверия и трех полукружных каналов. 1. Каналы и полости в височной кости образуют костный лабиринт вестибулярного аппарата, который частично заполнен перепончатым лабиринтом. Между костным и перепончатым лабиринтами находится жидкость – перилимфа, а внутри перепончатого лабиринта – эндолимфа.
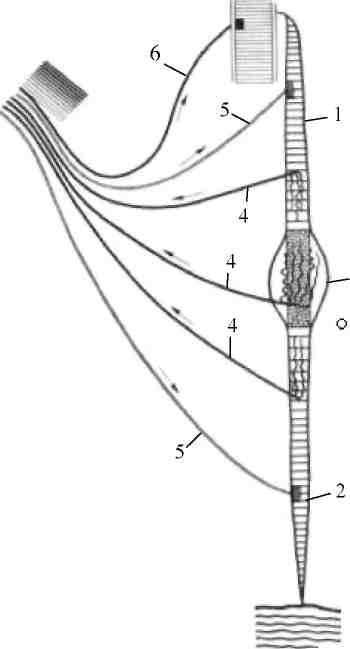
2. Аппарат преддверия предназначен для анализа действия силы тяжести при изменениях положения тела в пространстве и ускорений прямолинейного движения. Он разделен на 2 полости – мешочек и маточку, содержащих отолитовые приборы, механорецепторы которых представляют собой волосковые клетки. Выступающая в полость часть рецепторной клетки оканчивается одним более длинным подвижным волоском и 60 – 80 склеенными неподвижными волосками. Эти волоски пронизывают желеобразную отолитовую мембрану, в которой находятся кристаллы углекислого кальция – отолиты (рис. 33). 3. В маточке отолитовая мембрана расположена в горизонтальной плоскости, а в мешочке она согнута и находится во фронтальной и сагиттальной плоскостях. 4. При изменении положения головы и тела, а также при вертикальных или горизонтальных ускорениях отолитовые мембраны свободно перемещаются под действием силы тяжести во всех трех плоскостях (т.е. скользят по волоскам), деформируя при этом волоски механорецепторов. Чем больше деформация волосков, тем выше частота афферентных импульсов в волокнах вестибулярного нерва.
Рис. 33. Строение отолитового аппарата: 1 – отолиты; 2 – отолитовая мембрана; 3 – волоски рецепторных клеток; 4 – рецепторные клетки; 5 – опорные клетки; 6 – нервные волокна
Аппарат полукружных каналов служит для анализа действия центробежной силы при вращательных движениях. Адекватным его раздражителем является угловое ускорение. Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях (передняя – во фронтальной плоскости, боковая – в горизонтальной, задняя – в сагиттальной ) и заполнены, как и весь лабиринт, плотной эндолимфой (ее вязкость в 2 – 3 раза больше, чем у воды). Один из концов каждого канала расширен в «ампулу». Рецепторные волосковые клетки сконцентрированы только в ампулах в виде крист (складок, гребешков), т.е. склеены. При движении эндолимфы (во время угловых ускорений), когда волоски сгибаются в одну сторону, волосковые клетки возбуждаются, а при противоположно направленном движении – тормозятся. Рецепторный потенциал, генерируемый при раздражении волосковых клеток, передает импульс окончаниям волокон вестибулярного нерва. В настоящее время показано, что вращения или наклоны в одну сторону увеличивают афферентную импульсацию, а в другую сторону – уменьшают ее. Это позволяет различать направление прямолинейного или вращательного движения. 6. Влияние вестибулярной системы на различные функции организма Вестибулярная сенсорная система связана со многими центрами спинного и головного мозга и вызывает ряд вестибулосоматических и вестибуловегетативных рефлексов (рис. 34). Важнейшие из этих реакций – вестибулоспинальные. Вестибулярные раздражения вызывают установочные рефлексы изменения тонуса мышц, лифтные рефлексы, а также особые движения глаз, направленные на сохранение изображения на сетчатке, – нистагм (движения глазных яблок со скоростью вращения, но в противоположном направлении, затем быстрое возвращение к исходной позиции и новое противоположное вращение).
Рис. 34. Афферентные связи вестибулярного аппарата: Г – глаз; Тк – тонкая кишка; М – мышца; Пм – продолговатый мозг; Ж – желудок; См – спинной мозг В вестибуловегетативные реакции вовлекаются сердечнососудистая система, желудочно-кишечный тракт и другие органы. При сильных и длительных нагрузках на вестибулярный аппарат возникает «болезнь движения» (примером которой может служить морская болезнь), которая проявляется изменением частоты сердцебиения и артериального давления, ухудшением чувства времени, изменением психических функций – внимания, оперативного мышления, кратковременной памяти, эмоциональных проявлений. В тяжелых случаях возникают головокружение, тошнота, рвота. Повышенная склонность к «болезни движения» может быть уменьшена специальной тренировкой (вращение, качели) и применением ряда лекарственных средств. В условиях невесомости (когда у человека выключены вестибулярные влияния) возникает утрата представления о пространственном положении тела. Теряются навыки ходьбы, бега. Ухудшается состояние нервной системы, возникает повышенная раздражительность, нестабильность настроения. Таким образом, помимо основной анализаторной функции, важной для управления позой и движениями человека, вестибулярная сенсорная система оказывает разнообразные побочные влияния на многие функции организма, которые возникают в результате иррадиации возбуждения на другие нервные центры.
Структура Функция (выполняемая изолированной структурой)
Роль в осуществлении движения
Побуждение к действию Замысел действия Схемы целенаправленных движений (приобретенные и врожденные) Регуляция позы Моно- и полисинаптические рефлексы Длина мышц Напряжение мышц у л У J у J План Программа Выполнение Рис. 35. Общий план организации двигательной сенсорной системы Лекция 22 ДВИГАТЕЛЬНАЯ СЕНСОРНАЯ СИСТЕМА. СЕНСОРНЫЕ СИСТЕМЫ КОЖИ, ВКУСА И ОБОНЯНИЯ 1. Значение и общий план организации двигательной сенсорной системы Двигательная сенсорная система служит для анализа состояния двигательного аппарата – его движения и положения. Информация о степени сокращения скелетных мышц, натяжении сухожилий, изменении суставных углов необходима для регуляции двигательных актов и поз. Двигательная сенсорная система состоит из следующих отделов:
1. периферического, представленного проприорецепторами, расположенными в мышцах, сухожилиях и суставных сумках; 2. проводникового, который начинается биполярными клетками (первыми нейронами), тела которых расположены вне ЦНС – в спинномозговых узлах, один их отросток связан с рецепторами, другой входит в спинной мозг и передает импульсы ко вторым нейронам в продолговатый мозг (часть путей от проприорецепторов направляется в кору мозжечка), а далее к третьим нейронам – релейным ядрам таламуса; 3. коркового, находящегося в передней центральной извилине коры больших полушарий. Общий план организации двигательной сенсорной системы представлен на рис. 35. 2. Функции проприорецепторов В мышцах млекопитающих и человека содержатся 3 типа специализированных рецепторов: мышечные веретена, сухожильные рецепторы
Гольджи и суставные рецепторы (рецепторы суставной капсулы и суставных связок). Все эти рецепторы реагируют на механические раздражения и участвуют в координации движений, являясь источником информации о состоянии двигательного аппарата. Специфическим раздражителем проприорецепторов является их растяжение. Мышечные веретена представляют собой небольшие продолговатые образования (длиной несколько миллиметров, шириной - десятые доли миллиметра), расположенные в толще мышцы. Каждое веретено покрыто капсулой, образованной несколькими слоями клеток, которая в центральной части расширяется и образует ядерную сумку (рис. 36). Рис. 36. Мышечное веретено: 1 – проксимальный конец интрафузального мышечного волокна, прикрепленного к волокну скелетной мышцы; 2 – дистальный конец этого волокна, прикрепленного к фасции; 3 – ядерная сумка; 4 – афферентные волокна; 5 – волокна гамма-мотонейрона; 6 – волокно альфа-мотонейрона, идущее к скелетной мышце Внутри капсулы находится пучок (от 2 до 14) тонких волокон (в 2 - 3 раза тоньше обычных волокон скелетных мышц), которые называют интрафузальными в отличие от всех остальных волокон мышцы (экстрафузальных). Веретена расположены параллельно экстрафузальным волокнам -один конец прикреплен к сухожилию, а другой - к волокну. Различают ин-трафузальные волокна двух типов: ■ ядерносумчатые - более толстые и длинные с ядрами в средней, утолщенной, части волокна - ядерной сумке, которые связаны с наиболее толстыми и быстропроводящими афферентными нервными волокнами - они информируют о динамическом компоненте движения (скорости изменения длины мышцы); ■ ядерноцепочечные - более короткие, тонкие, с ядрами, вытянутыми в цепочку, информирующие о статическом компоненте (удерживаемой в данный момент длине мышцы). На интрафузальных волокнах спирально расположены (намотаны) чувствительные окончания афферентных нервных волокон. При растяжении скелетной мышцы происходит растяжение и мышечных рецепторов, при этом деформируются окончания нервных волокон, что вызывает появление в них нервных импульсов, идущих, в первую очередь, к мотонейронам спинного мозга. Частота импульсации возрастает с увеличением растяжения мышцы, а также при увеличении скорости ее растяжения. Тем самым нервные центры информируются о скорости растяжения мышцы и ее длине. Импульсация от мышечных веретен продолжается в течение всего периода поддержания растянутого состояния, что обеспечивает постоянную осведомленность центров о длине мышцы. Чем более тонкие и координированные движения осуществляют мышцы, тем больше в них мышечных веретен: у человека в глубоких мышцах шеи, связывающих позвоночник с головой, среднее их число составляет 63, а в мышцах бедра и таза - менее 5 веретен на 1 г веса мышцы.
ЦНС может тонко регулировать чувствительность проприорецепторов, т.е. веретена имеют и эфферентную иннервацию: интрафузальные мышечные волокна иннервируются аксонами, идущими к ним от гамма-мотонейронов. Возбуждение альфа-мотонейронов сопровождается возбуждением гамма-мотонейронов. Активация гамма-мотонейронов приводит к повышению чувствительности (возбудимости) афферентных нейронов: при той же длине скелетной мышцы в нервные центры при этом будет поступать большее число афферентных импульсов. Разряды мелких гамма-мотонейронов спинного мозга вызывают сокращение интрафузальных мышечных волокон по обе стороны от ядерной сумки веретена. В результате средняя несократимая часть мышечного веретена растягивается, и деформация отходящего отсюда нервного волокна вызывает повышение его возбудимости. Это позволяет, во-первых, выделять проприоцептивную импульсацию на фоне другой афферентной информации и, во-вторых, увеличивать точность анализа состояния мышц. Повышение чувствительности веретен происходит во время движения и даже в предстартовом состоянии. Это объясняется тем, что в силу низкой возбудимости гамма-мотонейронов их активность в состоянии покоя выражена слабо, а при произвольных движениях и вестибулярных реакциях она активируется. Чувствительность проприорецепторов повышается также при умеренных раздражениях симпатических волокон и выделении небольших доз адреналина. Сухожильные рецепторы Гольджи находятся в месте соединения мышечных волокон с сухожилием. Сухожильные рецепторы (окончания нервных волокон) оплетают тонкие сухожильные волокна, окруженные капсулой. В результате последовательного крепления сухожильных рецепторов к мышечным волокнам (а в ряде случаев – к мышечным веретенам) растяжение сухожильных механорецепторов происходит при напряжении мышц, т.е. они возбуждаются при сокращении мышцы. Таким образом, в отличие от мышечных веретен сухожильные рецепторы информируют нервные центры о силе, развиваемой мышцей (о степени напряжения мышц и скорости его развития). На спинальном уровне они через интернейроны вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов антагониста. Суставные рецепторы информируют о положении отдельных частей тела в пространстве и относительно друг друга. Они представляют собой свободные нервные окончания или окончания, заключенные в специальную капсулу. Одни суставные рецепторы посылают информацию о величине суставного угла, т.е. о положении сустава. Их импульсация продолжается в течение всего периода сохранения данного угла. Она тем большей частоты, чем больше сдвиг угла. Другие суставные рецепторы возбуждаются только в момент движения в суставе, т.е. посылают информацию о скорости движения. Частота их импульсации возрастает с увеличением скорости изменения суставного угла. Сигналы, идущие от рецепторов мышечных веретен, сухожильных органов, суставных сумок и тактильных рецепторов кожи, называют кинестетическими, т.е. информирующими о движении тела. Их участие в произвольной регуляции движений различно. Сигналы от суставных рецепторов вызывают заметную реакцию в коре больших полушарий и хорошо осознаются. Благодаря им человек лучше воспринимает различия при движениях в суставах, чем различия в степени напряжения мышц при статических положениях или поддержании веса. Сигналы же от других проприорецепторов, поступающие преимущественно в мозжечок, обеспечивают бессознательную регуляцию, подсознательный контроль движений и поз. 3. Сенсорные системы кожи, внутренних органов, вкуса и обоняния В коже и внутренних органах имеются разнообразные рецепторы, реагирующие на физические и химические раздражители. Кожная рецепция В коже представлена тактильная, температурная и болевая рецепция. На 1 см2 кожи, в среднем, приходится 12 13 холодовых точек, 1 2 тепловых, 25 тактильных и около 100 болевых. Тактильная сенсорная система предназначена для анализа давления и прикосновения. Ее рецепторы представляют собой свободные нервные окончания и сложные образования (тельца Мейснера, тельца Пачини), в которых нервные окончания заключены в специальную капсулу. Они находятся в верхних и нижних слоях кожи, в кожных сосудах, в основаниях волос. Особенно их много на пальцах рук и ног, ладонях, подошвах, губах. Это механорецепторы, реагирующие на растяжение, давление и вибрацию. Наиболее чувствительным рецептором является тельце Пачини, которое вызывает ощущение прикосновения при смещении капсулы лишь на 0,0001 мм. Чем больше размеры тельца Пачини, тем более толстые и быстропроводящие афферентные нервы отходят от него. Они проводят кратковременные залпы (длительностью 0,005 с), информирующие о начале и окончании действия механического раздражителя. Путь тактильной информации следующий: рецептор -► 1-й нейрон в спинномозговых узлах —► 2-й нейрон в спинном или продолговатом мозге —► 3- й нейрон в промежуточном мозге (в таламусе) —► 4- й нейрон в задней центральной извилине коры больших полушарий (в первичной соматосенсорной зоне). Температурная рецепция осуществляется холодовыми рецепторами (колбы Краузе) и тепловыми (тельца Руффини, Гольджи-Маццони). При температуре кожи 31 - 37 °С эти рецепторы почти неактивны. Ниже этой границы холодовые рецепторы активизируются пропорционально падению температуры, затем их активность падает и совсем прекращается при +12 °С. При температуре выше 37 °С активизируются тепловые рецепторы, достигая максимальной активности при +43 °С, затем резко прекращают ответы. Болевая рецепция, как считает большинство специалистов, не имеет специальных воспринимающих образований. Болевые раздражения воспринимаются свободными нервными окончаниями, а также возникают при сильных температурных и механических раздражениях в соответствующих термо- и механорецепторах. Температурные и болевые раздражения передаются в спинной мозг, оттуда в промежуточный мозг и в соматосенсорную область коры. 3.2. Висцероцептивная ( интерорецептивная ) сенсорная система Во внутренних органах имеется множество рецепторов, воспринимающих давление, – барорецепторы сосудов, кишечного тракта и др., изменения химизма внутренней среды, – хеморецепторы, ее температуры, – терморецепторы, осмотического давления, болевые раздражения. С их помощью безусловнорефлекторным путем регулируется постоянство различ-ных констант внутренней среды (поддержание гомеостаза), ЦНС информируется об изменениях во внутренних органах. Информация от интерорецепторов через блуждающий, чревный и тазовый нервы поступает в промежуточный мозг (и к таламусу, и к гипоталамусу), а также к подкорковым ядрам (хвостатому телу), мозжечку и далее – в лобные и другие области коры головного мозга. Деятельность этой системы практически не осознается, она мало локализована, однако при сильных раздражениях она хорошо ощущается. Она участвует в формировании сложных ощущений – жажды, голода и др. 3.3. Обонятельная и вкусовая сенсорные системы Обонятельная и вкусовая сенсорные системы относятся к древнейшим системам. Они предназначены для восприятия и анализа химических раздражений, поступающих из внешней среды. X еморецепторы обоняния находятся в обонятельном эпителии верхних носовых ходов. Это – волосковые биполярные клетки, передающие информацию через решетчатую кость черепа к клеткам обонятельной луковицы мозга и далее через обонятельный тракт к обонятельным зонам коры (крючок морского коня, извилина гиппокампа и другие). Различные рецепторы избирательно реагируют на разные молекулы пахучих веществ, возбуждаясь лишь теми молекулами, которые являются зеркальной копией поверхности рецептора. Они воспринимают эфирный, камфарный, мятный, мускусный и другие запахи, причем к некоторым веществам чувствительность необычайно высока. Хеморецепторы вкуса представляют собой вкусовые луковицы, расположенные в эпителии языка, задней стенке глотки и мягкого неба. У детей их количество больше, а с возрастом – убывает. Микроворсинки рецепторных клеток выступают из луковицы на поверхность языка и реагируют на растворенные в воде вещества. Их сигналы поступают через волокна лицевого и языко-глоточного нервов в таламус и далее в соматосенсорную область коры. Рецепторы разных частей языка воспринимают четыре основных вкуса: горький (задняя часть языка), кислый (края языка), сладкий (передняя часть языка) и соленый (передняя часть и края языка). Между вкусовыми ощущениями и химическим строением вещества отсутствует строгое соответствие, т.к. вкусовые ощущения могут изменяться при заболевании, беременности и т.д. В формировании вкусовых ощущений участвуют обоняние, тактильная, болевая и температурная чувствительность. Информация вкусовой сенсорной системы используется для организации пищевого поведения, связанного с добыванием, выбором, предпочтением или отверганием пищи, формированием чувства голода, сытости. 4. Переработка, взаимодействие и значение сенсорной информации Сенсорная информация передается от рецепторов в высшие отделы мозга по двум основным путям нервной системы – специфическим и неспецифическим. Специфические проводящие пути – это классические афферентные пути зрительной, слуховой, двигательной и других сенсорных систем, которые составляют один из трех основных функциональных блоков мозга – блок приема, переработки и хранения информации (А. Р. Лурия, 1962, 1973). В обработке этой информации участвует и неспецифическая система мозга, не имеющая прямых связей с периферическими рецепторами, но получающая импульсы по коллатералям от всех восходящих специфических систем и обеспечивающая их широкое взаимодействие. 4.1. Обработка сенсорной информации в проводниковых отделах Анализ получаемых раздражений происходит во всех отделах сенсорных систем. Наиболее простая форма анализа осуществляется уже на уровне рецепторов: из всех падающих на организм воздействий они выделяют (выбирают) раздражители одного вида (свет, звук и пр.). При этом в одной сенсорной системе возможно уже более детальное выделение характеристик сигналов ( цветоразличение фоторецепторами колбочек и др.). Дальнейшая обработка афферентной информации в проводниковом отделе заключается, с одной стороны, в продолжающемся анализе свойств раздражителя, а с другой – в процессах их синтеза, в обобщении поступившей информации. По мере передачи афферентных импульсов на более высокие уровни сенсорных систем возрастает сложность обработки информации: например, в подкорковых зрительных центрах среднего мозга есть нейроны, которые реагируют на различную степень освещенности и обнаруживают движение; в подкорковых слуховых центрах – нейроны, извлекающие информацию о высоте тона и локализации звука, что лежит в основе ориентировочного рефлекса на неожиданные раздражители, т.е. эти нейроны реагируют на афферентные сигналы более сложно, чем простые проводники. Благодаря многим разветвлениям афферентных путей на уровне спинного мозга и подкорковых центров обеспечивается многократное взаимодействие афферентных импульсов в пределах одной сенсорной системы, а также взаимодействие между различными сенсорными системами (в частности, можно отметить чрезвычайно обширные взаимодействия вестибулярной сенсорной системы со многими восходящими и нисходящими путями). Особенно широкие возможности для взаимодействия различных сигналов создаются в неспецифической системе мозга, где к одному и тому же нейрону могут сходиться (конвергировать) импульсы различного происхождения (от 30 тысяч нейронов) и от разных рецепторов тела. Вследствие этого неспецифическая система играет большую роль в процессах интеграции функций в организме. При поступлении в более высокие уровни ЦНС происходит либо сжатие, либо расширение информации, приходящей от одного рецептора, что связано с неодинаковым числом элементов в соседних слоях. Примером может служить зрительная сенсорная система, где слой фоторецепторов в каждой из двух сетчаток человека имеет около 130 млн элементов, а в слое выходных – ганглиозных клеток сетчатки – всего 1 млн 250 тысяч нейронов. Одна ганглиозная клетка сетчатки объединяет информацию от сотни биполярных клеток и десятков тысяч рецепторов, т.е. такая информация поступает в зрительные нервы уже после значительной обработки, в сокращенном виде. Это пример сужения (сжатия) информации. С другой стороны, сигналы одного рецептора связаны с десятками ганглиозных клеток и могут, в принципе, передавать информацию любым корковым нейронам зрительной коры. На более высоких уровнях зрительной сенсорной системы происходит расширение информации: число нейронов в первичной зрительной зоне коры в тысячи раз больше, чем в подкорковом зрительном центре или на выходе из сетчатки. В слуховой и ряде других сенсорных систем представлена только расширяющаяся «воронка» – по направлению от рецепторов к коре. Физиологический смысл расширяющихся «воронок» – обеспечение более дробного и сложного анализа сигналов. Большое количество параллельных каналов (в зрительном нерве 900 000, а слуховом – 30 000 волокон) обеспечивает передачу без искажений специфической информации от рецепторов к коре. Одной из важнейших сторон обработки афферентной информации является отбор наиболее значимых сигналов, осуществляемый восходящими и нисходящими влияниями на различных уровнях сенсорных систем. В этом отборе важную роль играет также неспецифический отдел нервной системы (лимбическая система, ретикулярная формация). Активируя или затормаживая многие центральные нейроны, он способствует отбору наиболее значимой для организма информации. В отличие от обширных влияний среднемозговой части ретикулярной формации, импульсация из неспецифических ядер таламуса воздействует лишь на ограниченные участки коры больших полушарий. Такое избирательное повышение активности небольшой территории коры имеет значение в организации акта внимания, выделяя на общем афферентном фоне наиболее важные в данный момент сообщения. 4.2. Обработка информации на корковом уровне В коре больших полушарий сложность обработки информации возрастает от первичных полей к вторичным и третичным ее полям. Первичные поля коры осуществляют анализ раздражений определенного вида, поступающих от связанных с ними специфических рецепторов. Это так называемые ядерные зоны анализаторов (по И. П. Павлову) – зрительные, слуховые и др. Их деятельность лежит в основе возникновения ощущений. Лежащие вокруг них вторичные поля (периферия анализаторов) получают от первичных полей результаты обработки информации и преобразуют их в более сложные формы. Во вторичных полях происходит осмысливание полученной информации, ее узнавание, обеспечиваются процессы восприятия раздражений данного вида. От вторичных полей отдельных сенсорных систем информация поступает в задние третичные поля – ассоциативные нижнетеменные зоны, где происходит интеграция сигналов различной модальности, позволяющая создать цельный образ внешнего мира со всеми его запахами, звуками, красками и т.п. Здесь на основе афферентных сообщений от разных частей правой и левой половины тела формируются сложные представления человека о схеме пространства и схеме тела, которые обеспечивают пространственную ориентацию движений и точную адресацию моторных команд к различным скелетным мышцам. Эти зоны также имеют особое значение в хранении полученной информации. На основе анализа и синтеза информации, обработанной в заднем третичном поле коры, в ее передних третичных полях (передней лобной области) формируются цели, задачи и программы поведения человека. Важной особенностью корковой организации сенсорных систем является экранное или соматотопическое (лат. соматикус – телесный, топикус – местный) представительство функций. Чувствительные корковые центры первичных полей коры образуют как бы экран, отражающий расположение рецепторов на периферии, т.е. здесь имеются проекции «точка в точку». Так, в задней центральной извилине (в соматосенсорной зоне) нейроны тактильной, температурной и кожной чувствительности представлены в том же порядке, что и рецепторы на поверхности тела, напоминая копию человечка (гомункулюса); в зрительной коре – как бы экран рецепторов сетчатки; в слуховой коре – в определенном порядке нейроны, реагирующие на определенную высоту звуков. Тот же принцип пространственного представительства информации наблюдается в переключательных ядрах таламуса, в коре мозжечка, что значительно облегчает взаимодействие различных отделов ЦНС. Область коркового сенсорного представительства по своим размерам отражает функциональную значимость той или иной части афферентной информации. Так, в связи с особой значимостью анализа информации от кинестетических рецепторов пальцев руки и от речеобразующего аппарата у человека территория их коркового представительства значительно превосходит сенсорное представительство других участков тела. Аналогично этому, на единицу площади центральной ямки в сетчатке глаза приходится почти в 500 раз большая зона зрительной коры, чем на такую же единицу площади периферии сетчатки. Высшие отделы ЦНС обеспечивают активный поиск сенсорной информации. Это наглядно проявляется в деятельности зрительной сенсорной системы. Специальные исследования движений глаз показали, что взор фиксирует не все точки пространства, а лишь наиболее информативные признаки, особо важные для решения какой - либо задачи в данный момент. Поисковая функция глаз является частью активного поведения человека во внешней среде, его сознательной деятельностью. Она управляется высшими анализирующими и интегрирующими областями коры – лобными долями, под контролем которых происходит активное восприятие внешнего мира.
Кора больших полушарий обеспечивает наиболее широкое взаимодействие различных сенсорных систем и их участие в организации двигательных действий человека, в т.ч. в процессе его спортивной деятельности. 4.3. Значение деятельности сенсорных систем в спорте Эффективность выполнения спортивных упражнений зависит от процессов восприятия и переработки сенсорной информации. Четкое восприятие пространства и пространственная ориентация движений обеспечиваются функционированием зрительной, слуховой, вестибулярной, кинестетической рецепции. Оценка временных интервалов и управление временными параметрами движений базируется на проприоцептивных и слуховых ощущениях. Вестибулярные раздражения при поворотах, вращениях, наклонах и т.п. заметно влияют на координацию движений и проявление физических качеств, особенно, при низкой устойчивости вестибулярного аппарата. Экспериментальное выключение отдельных сенсорных афферентаций у спортсменов (выполнение движений в специальном ошейнике, исключающем активацию шейных проприорецепторов; использование очков, закрывающих центральное или периферическое поле зрения ) приводило к резкому снижению оценок за упражнение или к полной невозможности его исполнения. В противоположность этому, сообщение спортсмену дополнительной информации (особенно срочной – в процессе движения) помогало быстрому совершенствованию технических действий. На основе взаимодействия сенсорных систем у спортсменов вырабатываются комплексные представления, сопровождающие его деятельность в избранном виде спорта – «чувство» льда, снега, воды и т.п. При этом в каждом виде спорта имеются наиболее важные – ведущие сенсорные системы, от активности которых в наибольшей мере зависит успешность выступлений спортсмена.
Материалы для самостоятельной подготовки Вопросы к коллоквиуму и для самоконтроля 1. Кем было создано учение об анализаторах? 2. Что называют анализатором? 3. Назовите общие принципы строения сенсорных систем. 4. В чем заключается принцип многослойности; многоканальности сенсорных систем? 5. На какие отделы делятся сенсорные системы? 6. Что такое рецепторы? Материалы для самостоятельной подготовки Вопросы к коллоквиуму и для самоконтроля 1 Кем было создано учение об анализаторах? 2 Что называют анализатором? 3 Назовите общие принципы строения сенсорных систем. 4 В чем заключается принцип многослойности; многоканальности сенсорных систем? 5 На какие отделы делятся сенсорные системы? 6 Что такое рецепторы? 7. Назовите основные функции сенсорных систем. 8. Какие виды рецепторов Вы знае
|
|||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-06; просмотров: 1931; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.138.120.251 (0.02 с.) |