Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Структурно-функциональное созревание нервных волоконСодержание книги
Похожие статьи вашей тематики
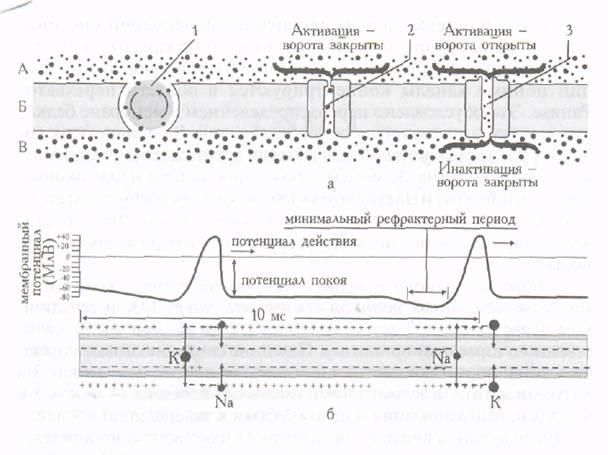
Поиск на нашем сайте Критерием структурно-функциональной зрелости мякотных и безмякотных нервных волокон является увеличение их толщины и уменьшение проницаемости клеточной мембраны, что оказывает существенное влияние на свойства нервного волокна. Однако главными критериями оценки зрелости мякотных нервных волокон являются их миелинизация, интенсивно происходящая к концу антенатального периода, и увеличение расстояния между перехватами Ранвье. Изменяется и распределение ионных каналов. У плода и ребенка первых лет жизни при неполной миелинизации нервных волокон натриевые и калиевые каналы в мембране распределяются равномерно. После завершения миелинизации ионные каналы концентрируются в области перехватов Ранвье. Это обусловлено перераспределением в мембране белковых молекул, являющихся основой каналов. В безмякотных волокнах распределение ионных каналов остается равномерным по всей длине волокна. К моменту рождения двигательные окончания в мышцах руки достигают более высокого уровня дифферен-цировки, чем в мышцах груди или спины, а также голени. Филогенетически старые пути миелинизируются раньше, чем филогенетически новые. У новорожденного в нервах голени, например, количество миелинизированных волокон составляет около 1/3, в седалищном нерве детей 2-3 лет - от 1/3 до 1/2 их общего числа, свойственного взрослому организму. Передние спинномозговые корешки у детей достигают состояния, присущего взрослым, между 2 и 5 годами жизни, а задние спинномозговые корешки - между 5 и 9 годами. Миелинизация в целом близка к завершению в 9 лет. Число аксонов в нерве с возрастом не изменяется, но в результате его созревания возбудимость, скорость проведения возбуждения и лабильность повышаются. Свойства нервных волокон Мембрана аксона разделяет жидкие среды, различающиеся по содержанию ионов натрия и калия (в жидкой среде, окружающей аксон, в 10 раз больше ионов натрия, чем ионов калия, внутри аксона - обратное соотношение). Встроенные в аксональную мембрану протеины играют роль селективных ионных каналов, пропускающих через мембрану ионы натрия или калия. В состоянии покоя, когда передача нервного импульса отсутствует, оба вида каналов закрыты и ионный (натрий-калиевый насос) поддерживает неравномерное (указанное выше) соотношение ионов натрия и калия, выкачивая наружу ионы натрия в обмен на ионы калия. Внутренняя поверхность мембраны в состоянии покоя заряжена отрицательно (приблизительно на 70 мВ) по отношению наружной мембране. При уменьшении заряда вследствие возникновения нервного импульса открываются натриевые каналы, пропускающие ионы натрия внутрь аксона. Вслед за этим натриевые каналы закрываются и открываются калиевые, выпускающие на наружную поверхность мембраны ионы калия. Ионные каналы последовательно открываются и закрываются, что обеспечивает распространение нервного импульса (рис.2).
Рис.2. Строение мембраны аксона (а) и распространение нервного импульса по аксону (б). Ионы натрия обозначены мелкими точками, ионы калия - крупными точками. А - наружная сторона мембраны аксона, Б - мембрана аксона, В - внутренняя сторона мембраны аксона. 1 - ионный насос, 2 - калиевый канал, 3 - натриевый канал
Распространению нервного импульса вдоль аксона способствует локализованное поступление внутрь аксона ионов натрия с последующим выходом ионов калия через каналы, которые открываются и закрываются под влиянием изменения разности потенциалов. Зарождение нервного импульса, который далее распространяется по аксону, происходит в теле нейрона. При небольшом изменении разности потенциалов открываются отдельные натриевые каналы, что еще больше увеличивает разность потенциалов. В месте поступления ионов натрия внутрь аксона внутренняя мембрана становится положительно заряженной. Возникающие изменения разности потенциалов приводят к тому, что натриевые каналы закрываются, открываются калиевые, калий выходит на наружную поверхность мембраны. Результатом этого процесса является быстрое восстановление отрицательного заряда внутренней мембраны. Изменение разности потенциалов носит название «потенциал действия». Потенциал действия распространяется по аксону. После короткого рефрактерного периода может возникать второй импульс. Возбудимость нервных волокон плода и новорожденного значительно ниже, чем у взрослого, но с 3-месячного возраста она начинает повышаться. Величина хронаксии в несколько раз больше, чем у взрослых. Потенциал покоя у детей значительно меньше, чем у взрослых, из-за большой проницаемости клеточной мембраны для ионов. В процессе созревания нервного волокна проницаемость его мембраны уменьшается, что ведет к возрастанию ионной асимметрии. Последняя и является непосредственной причиной увеличения потенциала покоя. При этом увеличивается и потенциал действия, что также свидетельствует о функциональной зрелости нервного волокна. Небольшая величина потенциала действия новорожденного сочетается с большой его продолжительностью и часто с отсутствием реверсии, что объясняется меньшей, чем у взрослого, ионной асимметрией. Фазовые изменения возбудимости во время возбуждения в раннем постнатальном онтогенезе также имеют особенности. В частности, длительность абсолютной рефлекторной фазы 5-8 мс, а относительной – 40-60 мс (у взрослых - 0,5-2 и 2 -10 мс соответственно). Проводимость нерва плода и детей низкая. У новорожденных, например, скорость проведения возбуждения по нервным волокнам не превышает 50 % скорости взрослых. Так, в локтевом нерве максимальная скорость составляет у взрослых 62 м/с, у новорожденных - 31 м/с. Плохо выражена изолированность проведения возбуждения. С возрастом скорость проведения возбуждения по нервным волокнам возрастает в результате их миелинизации, увеличения толщины нервного волокна и его потенциала действия. Миелинизация нервных волокон обеспечивает увеличение скорости проведения возбуждения вследствие того, что непрерывное проведение возбуждения сменяется сальтаторным и увеличивается расстояние между перехватами Ранвье (потенциал действия «перепрыгивает» на большее расстояние). Увеличение толщины безмякотного нервного волокна и еще не покрытого миелином мякотного волокна ведет к ускорению проведения возбуждения в результате уменьшения продольного сопротивления ионному току в аксоплазме. При увеличении же мембранного потенциала проведение возбуждения ускоряется, потому что большой потенциал действия быстрее вызывает возбуждение соседнего участка нервного волокна. Скорость распространения возбуждения по нервным волокнам у детей становится такой же, как у взрослых, к 5-9 годам, чтo связано с завершением миелинизации различных волокон в разные сроки и окончанием увеличения диаметров осевых цилиндров. Лабильность - функциональная подвижность нервной ткани (термин впервые сформулировал Н.Е.Введенский)- определяется максимальным ритмом возбуждения, который ткань может воспроизводить в соответствии с ритмом стимулов. Лабильность свидетельствует о скорости метаболических процессов в нервной ткани и обусловливает характер возбуждения и торможения в ней, лабильность нервного волокна у плода и детей первых лет жизни низкая. С возрастом она увеличивается: число потенциалов действия, которое способно воспроизвести волокно в 1 с у новорожденных, например, составляет 4-10, а у детей 5-9 лет приближается к норме взрослых (300-1000 импульсов).
|
||
|
|
Последнее изменение этой страницы: 2019-11-02; просмотров: 704; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.102 (0.01 с.) |