

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Тема 6. 3. Синтез гликогена (гликогеногенез), мобилизация гликогена (гликогенолиз). Регуляция процессов
Содержание книги
- Тема 2. 4. Кофакторы и коферменты
- Тема 2. 6. Основы кинетики ферментативного
- Тема 2. 7. Ингибиторы активности ферментов
- Тема 2. 8. Регуляция активности ферментов
- Тема 2. 9. Применение ферментов в медицине
- Модуль 3 матричные биосинтезы
- Тема 3. 2. Биосинтез днк (репликация)
- III. Исключение праймеров. Завершение формирования отстающей цепи ДНК
- Тема 3. 4. Биосинтез рнк (транскрипция). Посттранскрипционные модификации рнк
- Тема 3. 5. Трансляция как механизм перевода генетической информации в фенотипические
- Тема 3. 6. Ингибиторы матричных биосинтезов: лекарственные препараты, яды и бактериальные токсины
- Тема 3. 7. Механизмы адаптивной регуляции активности генов у прокариотов и эукариотов
- Тема 3. 8. Механизмы, обеспечивающие разнообразие белков у эукариотов
- Тема 3. 9. Механизмы генетической изменчивости: эволюционная изменчивость, полиморфизм белков. Наследственные болезни
- Тема 3. 10. Использование рекомбинантных днк
- Использование техники рекомбинанатных днк для диагностики и лечения заболеваний
- Инактивация аденилатциклазы и протеинкиназы А
- Последовательность событий передачи сигнала первичных мессенджеров с помощью инозитолфосфатной системы
- Первый этап тканевого дыхания - дегидрирование различных субстратов, образующихся в реакциях катаболизма.
- Тема 5. 4. Сопряжение тканевого дыхания и синтеза атф
- В реакциях цпэ часть энергии не превращается в энергию макроэргических связей атф, А рассеивается в виде теплоты.
- Окислительное декарбоксилирование пирувата
- Кроме того, адф аллостерически активирует некоторые ферменты опк
- Тема 5. 12. Гипоэнергетические состояния
- Желудочный сок не содержит ферментов, расщепляющих пищевые углеводы. Амилаза слюны инактивируется в желудке, так как оптимальное
- Тема 6. 3. Синтез гликогена (гликогеногенез), мобилизация гликогена (гликогенолиз). Регуляция процессов
- Активация гликогенфосфорилазы адреналином посредством аденилатциклазной системы
- В аэробном и анаэробном гликолизе можно выделить два этапа.
- Тема 6. 6. Биологическое значение катаболизма глюкозы. Регуляция процесса
- Анаболическое значение катаболизма глюкозы.
- Тема 6. 7. Пентозофосфатный путь превращения
- А - окислительный этап; б - неокслительный этап в обратном направлении
- Тема 6. 9. Регуляция гликолиза и глюконеогенеза
- Регуляция активности пируваткиназы в печени осуществляется путем фосфорилирования (дефосфорилирования) в зависимости от ритма питания
- Структурная организация межклеточного матрикса (суставной хрящ, базальные мембраны, субэпителиальные слои)
- Гликозилирование лизина под действием гликозилтрансфераз прекращается по мере формирования трехспиральной структуры.
- Патологий, связанных с уменьшением эластичности сосудов. При недостаточной активности металлопротеиназ развивается фиброз тканей и неадекватный иммунный ответ.
- Хондроитинсульфат; 2 - кератансульфат; 3 - коровый белок; гк - гиалуроновая кислота
- Тема 7. 5. Структурная организация межклеточного матрикса (суставной хрящ, базальные мембраны, субэпителиальные слои)
- Регуляция процесса. Амф, гмф, имф, ди- и трифосфаты адениловых и гуаниловых нуклеотидов ингибируют ключевые реакции своего синтезааллостерически по механизму отрицательной обратной связи.
- Частым нарушением катаболизма пуринов является гиперурикемия, которая возникает, когда в плазме крови Концентрация мочевой кислоты превышает норму.
- Синтез цтф из утф осуществляет цтф-синтетаза, используя амидную группу глн и энергию атф для аминирования пиримидинового кольца.
- Тема 10. 3. Биосинтез дезоксирибонуклеотидов.
- Тема 10. 4. Механизмы действия противовирусных и противоопухолевых препаратов на ферменты синтеза рибо- и дезоксирибонуклеотидов
- И рецепторов. Особенно много сфинголипидов в нервной ткани, где они формируют миелиновые оболочки нейронов.
- Тема 8. 3. Хиломикроны - транспортная форма экзогенных жиров
- Модульная единица 2 биосинтез высших жирных кислот и жиров
- Реакции восстановления обеспечивают синтез насыщенного алифатического радикала жирных кислот
- Запасание жиров в жировой ткани - так называемое депонирование жиров - происходит в абсорбтивный период, когда увеличивается соотношение инсулин - глюкагон.
- Первичное ожирение развивается в результате алиментарного дисбаланса - избыточной калорийности питания по сравнению с расходами энергии.
Похожие статьи вашей тематики
1. Гликоген - основной резервный полисахарид в клетках животных. Гликоген представляет собой разветвленный гомополисахарид, мономером которого является глюкоза. Остатки глюкозы соединены в линейных участках а1,4-гликозидными связями, а в местах разветвления - связями α1,6. Молекула гликогена более разветвлена, чем молекула крахмала, точки ветвления встречаются через каждые 8-10 остатков глюкозы. Разветвленная структура гликогена обеспечивает большое количество концевых мономеров, что способствует работе ферментов, отщепляющих или присоединяющих мономеры, так как эти ферменты могут одновременно работать на многих ветвях молекулы гликогена.
Гликоген депонируется главным образом в печени и скелетных мышцах и хранится в цитозоле клеток в форме гранул. Гранулы гликогена плохо растворимы в воде и не влияют на осмотическое давление в клетке. Это обстоятельство объясняет, почему в клетке депонируется гликоген, а не свободная глюкоза. С гранулами связаны и некоторые ферменты, участвующие в обмене гликогена, что облегчает взаимодействие ферментов с субстратами.
2. Синтез гликогена. Гликоген синтезируется в период пищеварения (абсорбтивный период: 1-2 часа после приема углеводной пищи) в основном в печени и в мышцах. Этот процесс требует затрат энергии, так включение одного мономера в полисахаридную цепь сопряжено с расходованием АТФ и УТФ (рис. 6.4, реакции 1 и 3). Образованная УДФ-глюкоза (реакция 3) является субстратом для гликогенсинтазы, которая переносит остаток глюкозы (реакция 4) на праймер (олигосахарид из 4-8 остатков глюкозы) и соединяет его а1,4-глюкозной связью. Когда длина синтезируемой цепи увеличивается на 11-12 остатков глюкозы, фермент ветвления - глюкозил- 1,4-1,6-трансфераза (реакция 5) образует боковую цепь путем переноса фрагмента из 5-6 остатков глюкозы на внутренний остаток глюкозы, соединяя
  его а1,6-гликозидной связью. Затем удлинение цепей и ветвление их повторяется много раз. В итоге образуется сильно разветвленная молекула, содержащая до 1млн глюкозных остатков. его а1,6-гликозидной связью. Затем удлинение цепей и ветвление их повторяется много раз. В итоге образуется сильно разветвленная молекула, содержащая до 1млн глюкозных остатков.
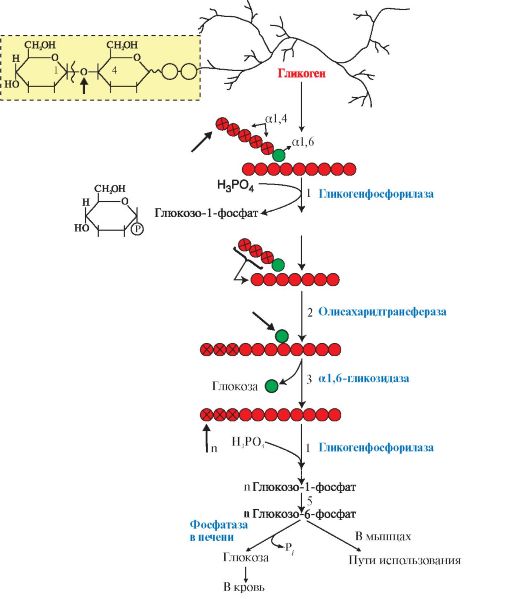
3. Мобилизация (распад) гликогена происходит в интервалах между приемами пищи (постабсорбтивный период) и ускоряется во время физической работы. Этот процесс осуществляется путем последовательного отщепления остатков глюкозы, в виде глюкозо-1-фосфата (рис. 6.5, реакция 1) с помощью гликогенфосфорилазы, расщепляющей а1,4-гликозидные связи. Этот фермент не расщепляет а1,6-гликозидные связи в местах разветвлений, поэтому необходимы еще два фермента, после действия которых остаток глюкозы в точке ветвления освобождается в форме свободной глюкозы (рис. 6.5, реакции 2 и 3). Гликоген распадается до глюкозо-6-фосфата и свободной глюкозы без затрат АТФ.
Мобилизация гликогена в печени отличается от таковой в мышцах одной реакцией (реакция 5), обусловленной наличием в печени фермента глюкозо- 6-фосфатазы.
Присутствие в печени глюкозо-6-фосфатазы обеспечивает главную функцию гликогена печени - высвобождение глюкозы в кровь в интервалах между едой для использования ее другими органами. Таким образом, мобилизация гликогена печени обеспечивает поддержание глюкозы в крови на постоянном уровне 3,3-5,5 ммоль в постабсорбтивном периоде. Это обстоятельство является обязательным условием для работы других органов и особенно мозга. Через 10-18 часов после приема пищи запасы гликогена в печени значительно истощаются, а голодание в течение 24 часов приводит к полному его исчерпанию.
Функция мышечного гликогена заключается в обеспечении клеток глю- козо-6-фосфатом, используемым в самой мышце для окисления и получения энергии или других целей (табл. 6.2).
Таблица 6.2. Особенности мобилизации гликогена в печени и в мышцах
   4. Переключение процессов синтеза и мобилизации гликогена в печени и мышцах происходит при переходе из абсорбтивного состояния в постабсорбтивное и из состояния покоя в режим физической работы. В переключении этих метаболических путей в печени участвуют инсулин, глюкагон и адреналин, а в мышцах - инсулин и адреналин. 4. Переключение процессов синтеза и мобилизации гликогена в печени и мышцах происходит при переходе из абсорбтивного состояния в постабсорбтивное и из состояния покоя в режим физической работы. В переключении этих метаболических путей в печени участвуют инсулин, глюкагон и адреналин, а в мышцах - инсулин и адреналин.
Влияние этих гормонов на синтез и распад гликогена осуществляется путем изменения в противоположном направлении активности двух ключевых ферментов: гликогенсинтазы и гликогенфосфорилазы с помощью их фосфорилирования и дефосфорилирования (рис. 6.6).
  Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы в крови. Инсулин и глюкагон постоянно присутствуют в крови, но при переходе из абсорбтивного периода в постабсорбтивный изменяется их относительная концентрация. Отношение концентраций инсулина и глюкагона в крови называютинсулин-глюкагоновым индексом, в зависимости от которого изменяется направление метаболизма гликогена в печени. Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы в крови. Инсулин и глюкагон постоянно присутствуют в крови, но при переходе из абсорбтивного периода в постабсорбтивный изменяется их относительная концентрация. Отношение концентраций инсулина и глюкагона в крови называютинсулин-глюкагоновым индексом, в зависимости от которого изменяется направление метаболизма гликогена в печени.
|




