
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Модуль 3 матричные биосинтезы
Модульная единица 1 БИОСИНТЕЗ ДНК И РНК. РЕПАРАЦИЯ ОШИБОК И ПОВРЕЖДЕНИЙ ДНК ТЕМА 3.1. СТРОЕНИЕ И ФУНКЦИИ ДНК И РНК 1. ДНК и РНК представляют собой линейные полимеры, построенные из нуклеотидов. Каждый нуклеотид состоит из трех компонентов: азотистого основания, являющегося производным пурина или пиримидина, пентозы (рибозы или дезоксирибозы) и остатка фосфорной кислоты. В состав любой из нуклеиновых кислот входят два производных пурина (аденин, гуанин) и два производных пиримидина: в РНК - цитозин и урацил, а в ДНК - цитозин и тимин (табл. 3.1 и 3.2). Таблица 3.1. Строение нуклеотидов РНК
Таблица 3.2. Строение нуклеотидов ДНК
Первичная структура нуклеиновых кислот (НК) - это порядок чередования нуклеотидов в полинуклеотидной цепи, связанных между собой 3',5'-фосфодиэфирной связью. Образующиеся полимеры имеют фосфатный остаток на 5'-конце и свободную -ОН-группу пентозы на 3'-конце (рис. 3.1). Штрихами обозначают углеродные атомы пентозы для того, чтобы отличать их от атомов, входящих в азотистые основания.
Для краткого изображения последовательности нуклеотидов в нуклеиновых кислотах пользуются однобуквенным кодом. При этом запись осуществляют слева направо таким образом, что первый нуклеотид имеет свободный 5'-фосфатный конец, а последний -ОН-группу в 3'-положении рибозы или дезоксирибозы.
Рис. 3.1. Первичная структура нуклеиновых кислот. Х = Н для ДНК, Х = ОН для РНК Связи в молекуле нуклеиновых кислот: 1 - 5'-фосфоэфирная; 2 - N-гликозидная; 3 - 3',5'-фосфодиэфирная 2. Пространственная структура ДНК. Вторичная структура представляет собой правозакрученную спираль (рис. 3.2), в которой две полинуклеотидные цепи расположены антипараллельно и удерживаются относительно друг друга за счет водородных связей между комплементарными азотистыми основаниями: А = Т и G = С. Цепи молекулы ДНК не идентичны, но комплементарны друг другу: если известна первичная структура одной цепи, то последовательность нуклеотидов другой цепи задается правилом комплементарности оснований: Т одной цепи соответствует А, а С - G в другой цепи. Поэтому в молекуле ДНК количество адениловых нуклеотидов равно количеству тимидиловых нуклеотидов (А = Т), а количество гуаниловых равно количеству цитидиловых нуклеотидов (G = С). Соотношение А + Т / G + С - величина постоянная и является видоспецифической характеристикой организма. Основания нуклеотидов обращены внутрь молекулы и лежат в одной плоскости, которая практически перпендикулярна оси спирали. Между основаниями, расположенными друг под другом, возникают гидрофобные взаимодействия. Дезоксирибозофосфатные остатки образуют остов спирали. На один виток спирали приходится 10 нуклеотидных пар. Третичная структура ДНК формируется в результате ее взаимодействия с белками. Каждая молекула ДНК упакована в отдельную хромосому, в составе которой разнообразные белки связываются с отдельными участками ДНК и обеспечивают суперспирализацию и компактизацию молекулы. В период покоя комплексы ДНК с белками распределены равномерно по объему ядра, образуя хроматин. Белки хроматина включают две группы: гистоны и негистоновые белки.
Гистоны - небольшие белки с молекулярной массой от 11 000 до 22 000 Д и высоким содержанием лизина и аргинина. Четыре типа гистонов в количестве восьми молекул (по две каждого вида) образуют комплекс - нуклеосомный кор. Этот комплекс за счет ионных связей взаимодействует с отрицательно заряженными фосфатными группами участка ДНК длиной около 146 нуклеотидных пар (примерно 1,75 витка вокруг кора) и образует структуру, называемую нуклеосомой. Между нуклеосома- ми находятся участки ДНК длиной около 30 нуклеотидных пар - линкерные участки, к которым присоединяются молекулы гистона Н1 (рис. 3.3). Негистоновые белки представлены множеством ферментов и белков, участвующих в синтезе ДНК, РНК, регуляции этих процессов и компактизации ДНК. 3. Пространственная структура РНК. В клетке существует три вида РНК: рибосомная (рРНК), транспортная (тРНК) и матричная (мРНК), каждая из которых выполняет свою особую функцию в синтезе белка. Вторичная структура РНК формируется в результате спирализации отдельных участков одноцепочечной РНК. В спирализованных участках, или шпильках, между комплементарными парами азотистых оснований А и U, G и С возникают водородные связи. Двуцепочечные фрагменты чередуются с неспирализованными участками молекулы, образующими петли.
Аденин и тимин связывают две, а гуанин и цитозин - три водородные связи
Восемь молекул гистонов четырех видов (Н2А, Н2В, Н3 и Н4) составляют нуклеосомный кор (ядро), на который наматывается ДНК, образуя примерно два витка Третичная структура РНК образуется за счет дополнительных водородных связей между нуклеотидами неспирализованных участков, полинуклеотидной цепью рРНК или мРНК и белками, обеспечивает дополнительную компактизацию и стабилизацию пространственной структуры молекулы. 4. Структура молекул ДНК и РНК является видоспецифической характеристикой организмов. Об этом свидетельствуют эксперименты по гибридизации ДНК-ДНК и ДНК-РНК. При нагревании до 80-90°С нуклеиновые кислоты денатурируют с разрушением пространственной структуры, образуя одноцепочечные молекулы. При медленном охлаждении такие молекулы способны восстанавливать двойную спираль и либо приобретать исходную структуру, либо образовывать гибриды: совершенные или несовершенные в зависимости от комплементарности нитей друг другу по всей длине цепей. На этой способности двух образцов ДНК или ДНК и РНК образовывать гибриды в процессе охлаждения после денатурации основан метод молекулярной гибридизации. Этот метод позволил установить следующие закономерности: • ДНК всех клеток одного организма идентична, а ДНК разных организмов одного вида обнаруживает очень высокое сходство, обеспечивая образование «совершенных гибридов»; • ДНК специфична для каждого вида и чем больше филогенетическая дистанция между видами, тем больше различий в строении принадлежащих им ДНК; • ДНК, выделенная из тканей определенного организма, содержит информацию о структуре всех видов РНК данного организма.
А - гибридизация ДНК-ДНК; Б - гибридизация ДНК-РНК
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 411; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.17.5.68 (0.006 с.) |

 Рис. 3.2. Двойная спираль ДНК.
Рис. 3.2. Двойная спираль ДНК. Рис. 3.3. Структура нуклеосом.
Рис. 3.3. Структура нуклеосом.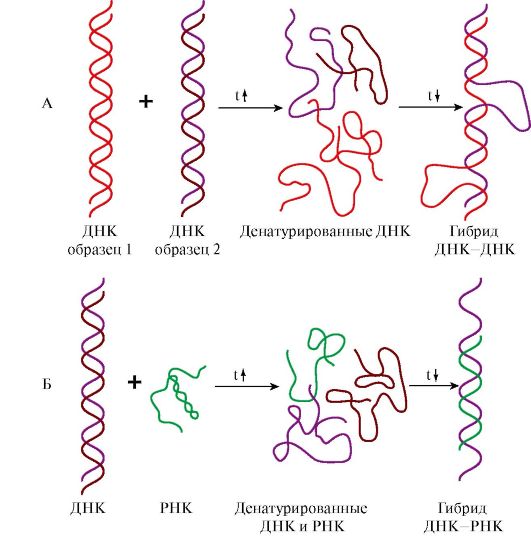 Рис. 3.4. Гибридизация нуклеиновых кислот:
Рис. 3.4. Гибридизация нуклеиновых кислот:


