Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
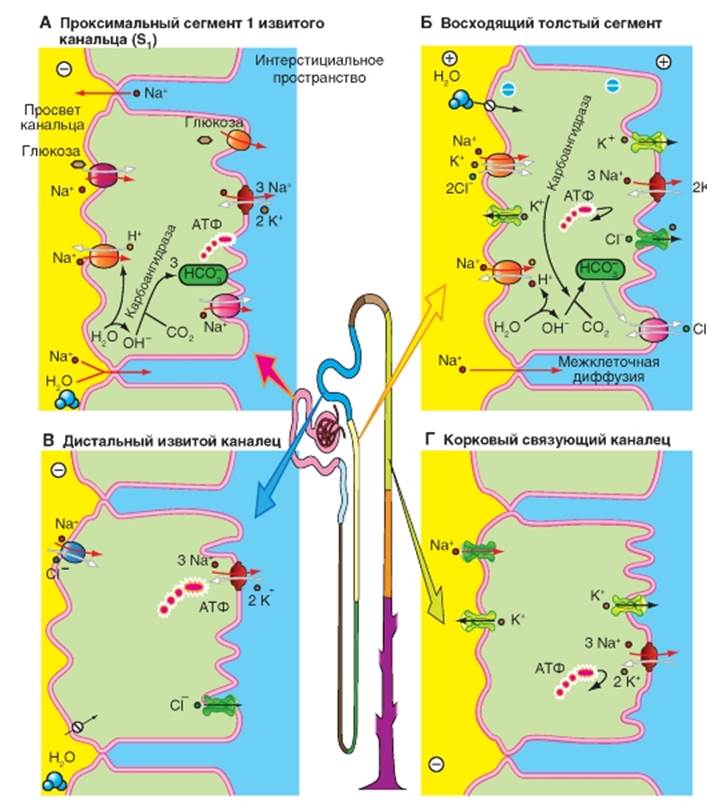
А - в проксимальном извитом канальце. Б - в дистальном прямом канальце (толстой восходящей части петли Генле). В - в дистальном извитом канальце. Г - в корковом связующем канальцеСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
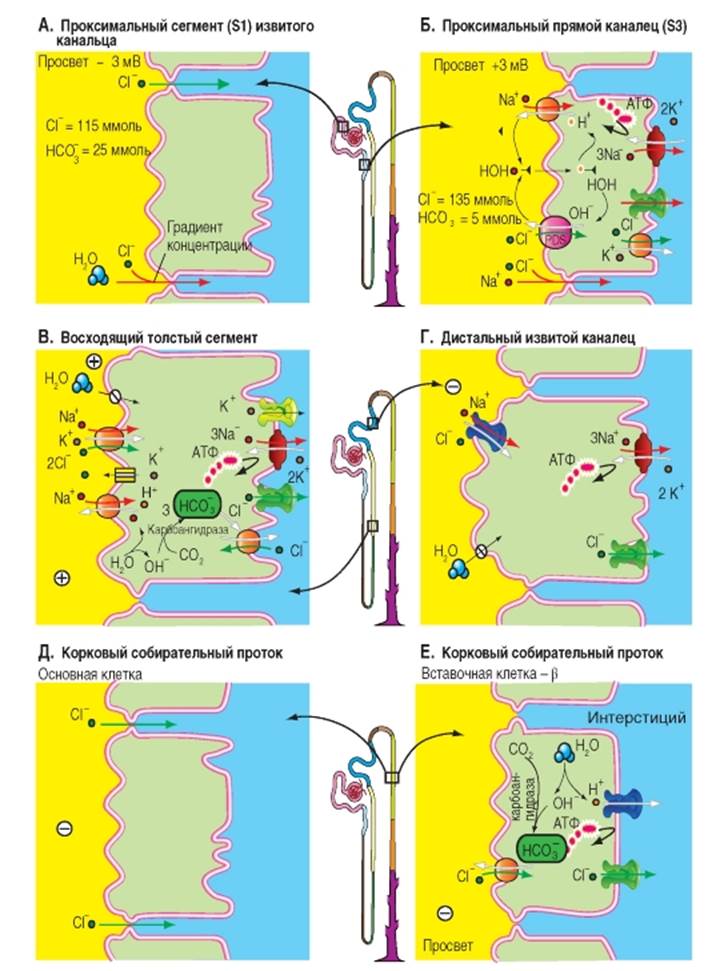
Реабсорбция Cl- в различных отделах нефрона В проксимальном извитом канальце Cl- реабсорбируется преимущественно межклеточно (рис. 9-11 А). В начальных отделах проксимального канальца (S1), где концентрация Cl- равна 115 ммоль, реабсорбция Cl- идет только вслед за водой (поток воды увлекает за собой растворенные в ней вещества: перенос вместе с растворителем или solvent drag). По мере продвижения фильтрата по канальцам несмотря на небольшую реабсорбцию Cl- его концентрация возрастает поскольку из просвета канальца уходит вода и Na+. Из-за реабсорбции воды концентрация Cl- в просвете канальца достигает 135 ммоль, то есть становится больше, чем концентрация Cl- в интерстициальной жидкости, (например, в просвете проксимального прямого канальца). Разница концентраций Cl- в просвете проксимального канальца по сравнению с концентрацией Cl- в интерстициальной жидкости на каждом участке канальца представляет собой движущую силу для межклеточной диффузии Cl- из просвета канальца в направлении к кровеносным сосудам. Таким образом, Cl-может покинуть просвет канальца под воздействием химической движущей силы (∆[Cl-]): через плотные контакты между апикальными участками мембраны эпителиальных клеток (межклеточная диффузия). Таким способом реабсорбируется часть профильтровавшегося Cl-. В результате этой диффузии Cl- далее по ходу проксимального канальца возникает трансэпителиальный потенциал, при котором жидкость просвета канальца несет положительный заряд (изменение знака потенциала), что в свою очередь обеспечивает межклеточную реабсорбцию катионов Na+, К+, Ca2+ и Mg2+. Величина трансэпителиального потенциала составляет 2 мВ. В проксимальном прямом канальце Cl- реабсорбируется как межклеточно вместе с Na+, так и при помощи переносчика (рис. 9-11 Б). В люминальной мембране доказано наличие переносчика (пендрин = PDS), который обменивает Cl- в том числе на формиат- (HCOO-). Кроме того клеточная мембрана проницаема (неионная диффузия) для муравьиной кислоты (HCOOH). При посредничестве переносчика NHE3 (антипорт Na+/H+), активность которого способствует циркуляции формиат- (НСОО-) через люминальную мембрану, Cl- накапливается в клетке результате вторичноактивного транспорта. Для этого, среда в просвете канальца должна быть более кислой чем в клетке, чтобы выделяемый формиат- (HCOO-) мог титроваться в HCOOH. Через базолатеральную мембрану Cl- выходит из клетки в интерстициум через Cl--каналы и с помощью К+/Cl--переносчи- ка (котранспорт). На базолатеральной мембране при этом идет активная реабсорбция Na+, при участии Na+/К+-АТФазы. Наиболее вероятный механизм для реабсорбции Na+ в средних и дальних участках проксимального канальца - обмен Na+/H+ (NНЕ3-переносчик), спаренный одновременно с обменом Cl-/OH- (PDS-переносчик). При этом OH- и Н+ взаимодействуют друг с другом в просвете канальца, превращаясь в воду, и в результате Na+ и Cl- входят в клетку эквимолярно. Из чего следует, что реабсорбция Cl- на этом участке в проксимального канальца осуществляется трансклеточно. Обнаружены еще два других механизма проникновения Cl- в клетку, в которые вовлечены процессы вторичного использов-ания формиата- (НСОО-) и секреции оксалата2 соответственно. Обмен Cl-/формиат- осуществляется PDS-переносчиком. Cl- выходит из клетки через базолатеральную мембрану пассивно через Cl--каналы, в результате осуществляемого белкомпереносчиком совместного транспорта К+-Cl- и в результате обмена Cl-/HCO3-. В дистальном прямом канальце (толстой восходящей части петли Генле) процесс реабсорбции осуществляется с помощью белка-переносчика (вторичный активный транспорт, котранспорт), расположенного на апикальной мембране клеток эпителия. Он переносит одновременно 1Na+, 1К+ и 2Cl-, при этом движущей силой является градиент Na+ опять создаваемый Na+/К+-АТФазой, расположенной на базолатеральной мембране (первичный активный транспорт, рис. 10 В). В результате вторично-активного транспорта, Cl-накапливается в клетках и покидает их через Cl--каналы типа CLC-Кb на базолатеральной мембране (рис. 9-11 В). Медикаментозно Na+- К+-2Cl--переносчик (NKCC2), расположенный со стороны просвета толстого восходящего отдела петли Генле, может быть ингибирован диуретиками (например, буметанидом). Поэтому данный переносчик называется также BSC1 (bumetanidsensitive cotransporter). Этот переносчик может также вместо Cl- переносить в клетку ионы NH4+. Реабсорбция в дистальном прямом канальце характеризуется высокой активностью процессов реабсорбции Na+. Со стороны просвета нефрона градиент Na+ используется для вторично-активного электронейтрального переноса Cl- и К+ (1). Через каналы К+ диффундирует обратно в просвет канальца (рециркуляция), а Cl- - в сторону кровеносных сосудов (2). Диффузия К+ из клетки приводит к гиперполяризации люминальной мембраны. Кроме того, наличие ионов К+ в просвете канальца устанавливает трансэпителиальный потенциал, при котором жидкость в просвете канальца по отношению к интерстициальной жидкости заряжена положительно (3). Под влиянием этого трансэпителияльного потенциала Na+, К+, Ca2+ и Mg2+ могут быть реабсорбированы пассивно через проницаемые для катионов плотные контакты (3). В конечном итоге, здесь активно реабсорбируется NaCl, за которым пассивно следуют катионы (включая Na+) (4). Трансмембранный перенос К+ через базолатеральные мембраны канальцев в интерстициум (который электрически нейтрализует выход Cl-) и перенос К+посредством Na+/К+-АТФазы здесь не изображены, чтобы не загромождать рисунок. Для воды этот сегмент канальца непроницаем, поэтому активный транспорт NaCl вызывает появление трансэпителиального осмотического градиента. В дистальном извитом канальце продолжается реабсорбция Na+ и Cl-, которые, как важные компоненты осмоляльности, все больше заменяются мочевиной, поставляемой из мозгового вещества почек (рис. 9-11 Г). NaCl из просвета канальца попадает в клетки дистального извитого канальца посредством механизма вторичного активного транспорта, обуславливающий одновременный перенос Na+ и Cl- (котранспорт; белок-переносчик: TSC), а в главные клетки связующего отдела и собирательной трубки - через Na+-каналы. Проникновение Na+ в клетку через Na+ -каналы деполяризует люминальную мембрану, что усиливает секрецию К+и способствует возникновению трансэпителиального потенциала, отрицательного в просвете канальца, под действием которого межклеточно реабсорбируется Cl-. В дистальном извитом канальце NaCl входит в клетку через апикальную мембрану посредством локализованного на люминальной мембране переносчика Na+ и Cl- (котранспорт), при этом Na+/ К+-АТФаза на базолатеральной мембране активно выводит Na+ из клетки, поддерживая электрохимический градиент, обеспечивающий вход Na+ через люминальную мембрану. Работа этого электронейтрального Na+-Cl--переносчика стимулируется альдостероном, и тормозится диуретиком тиацидом. Поэтому он был назван TSC (thiazidsensitive co-transporter). Cl- выходит из клетки через Cl--каналы (тип CLC-Кb). В корковом собирательном протоке (рис. 9-11 Д, Е) NaCl может реабсорбироваться против химического градиента, поэтому концентрация NaCl в конечной моче может снижаться в случае необходимости до нескольких ммоль. Рис.9-11. Клеточные модели транспорта Cl- в различных отделах нефрона. А - реабсорбция Cl- в проксимальном извитом канальце. Б - реабсорбция Cl- в проксимальном прямом канальце. В - реабсорбция Cl- в дистальном прямом канальце (толстой восходящей части петли Генле), Г - реабсорбция Cl- в дистальном извитом канальце, Д - реабсорбция Cl- в корковом собирательном протоке. Е - реабсорбция Cl- в β -вставочной клетке коркового собирательного протока
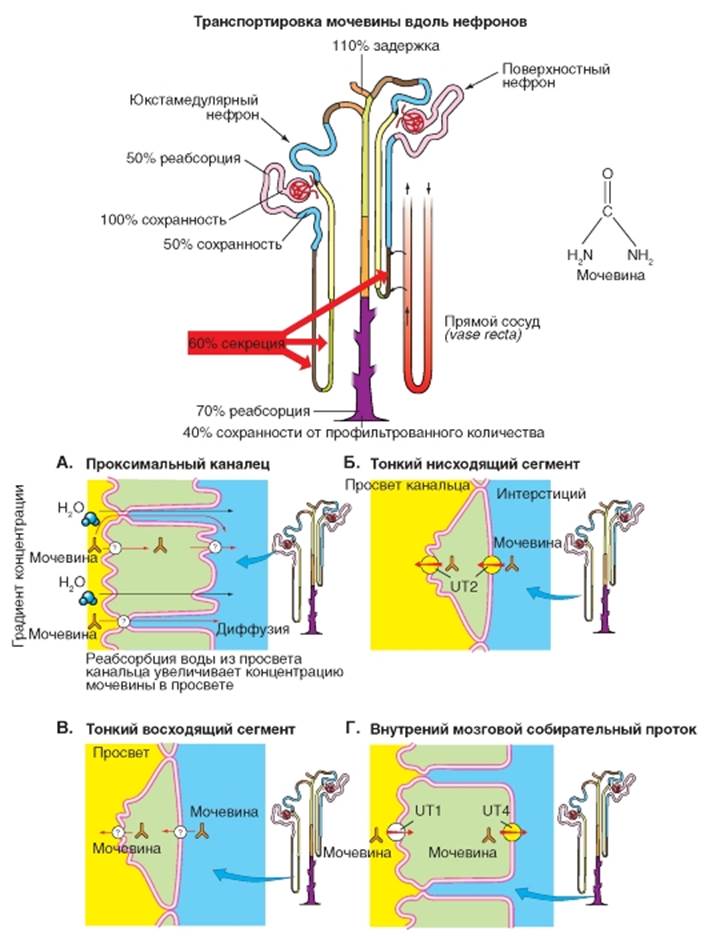
Транспорт мочевины До конца проксимального канальца реабсорбируется (пассивно) около половины профильтровавшейся мочевины, так как этот отдел нефрона проницаем для нее (рис. 9-12 А). Поскольку петля Генле погружается в богатый мочевиной интерстициум мозгового вещества почек, мочевина транспортируется из интерстициума в просвет тонкой нисходящей части петли Генле (рис. 9-12 Б), в стенке которой имеется переносчик мочевины (UT2 = Urea Transportert, Typ 2). В тонкой восходящей части петли Генле мочевина идет пассивно (диффузия) по градиенту концентрации. Концентрация мочевины в интерстициальной жидкости превышает ее концентрацию в просвете тонкой восходящей части петли Генле. Другие участки нефрона (толстая восходящая часть петли Генле, дистальный каналец, а также корковая и внешняя медуллярная собирательная трубочка) непроницаемы для мочевины, поэтому ее концентрация в просвете нефрона возрастает вследствие реабсорбции воды в этих отделах. В этих отделах нефрона мочевина даже замещает NaCl - важнейший компонент осмоляльности мочи. Лишь стенки нижних отделов собирательной трубочки вновь (в особенности в присутствии АДГ) проницаемы для мочевины. Там на апикальной мембране имеется переносчик UT1 (Urea Transportert, Typ 1), а на базолатеральной мембране - переносчик UT4 (Urea Transportert, Typ 4). При помощи эих двух переносчиков мочевина переносится из просвета собирательной трубочки по направлению ее химического градиента в клетку, а из нее в интерстициум внутреннего мозгового вещества (рис. 9-12 В). В конечном итоге происходит циркуляция мочевины между тонким нисходящим и восходящим отделом петли Генле и отделом собирательной трубочки, лежащей глубоко в мозговом веществе почки. Из медуллярного отдела собирательной трубочки вследствие ее высокой проницаемости для мочевины, в особенности в присутствии антидиуретического гормона, мочевина по градиенту концентрации пассивно выходит в мозговое вещество почки. Ее концентрация в интерстициальной жидкости превышает ее концентрацию в нисходящем тонком и восходящем отделах петли Генле, которые проницаемы для мочевины. Следующие отделы нефрона - толстая часть петли Генле, дистальный каналец и большая часть собирательной трубочки не проницаемы для мочевины и она не может покинуть эти отделы нефрона. В тех отделах нефрона (где в отличие от толстой восходящей части петли Генле нет активной реабсорбции NaCl: нисходящих и восходящих тонких сегментах петли Генле) мочевина принимает участие в процессе концентрирования мочи, до тех пор пока она в конце концов не покинет организм, как вещество, выводимое исключительно с мочой. Поскольку концентрация мочевины в интерстициуме мозгового вещества почки вблизи сосочка велика, она значительно влияет на общую осмоляльность, и поэтому концентрация NaCl в инерстициальной жидкости может поддерживаться на более низком уровне. Это способствует пассивной межклеточной реабсорбции NaCl из тонкой восходящей части петли Генле. Рис. 9-12. Транспорт мочевины. Мочевина является продуктом преобразования белков. А - проксимальный каналец. Б - тонкий нисходящий сегмент (нисходящая часть петли Генле). В - тонкий восходящий сегмент (восходящая часть петли Генле) и Г - внутренний мозговой собирательный проток проницаемы для мочевины, что приводит к ее частичной реабсорбции путем пассивной диффузии по градиенту концентрации. Прежде чем мочевина покинет почку, ее осмотическая активность используется для процесса концентрирования и для экономии Na+, так как некоторые отделы нефрона непроницаемы для мочевины. С начала дистального прямого канальца (толстой восходящей части петли Генле), в дистальном канальце и до начала нижней собирательной трубочки мочевина не может покинуть просвет нефрона, поэтому концентрация мочевины в просвете нефрона возрастает за счет оттока воды в дистальном извитом канальце и собирательной трубочке. Лишь во внутреннем медуллярном сегменте собирательной трубочки, особенно в присутствии AДГ мочевина переходит в интерстициум, где она в значительной мере определяет высокую осмоляльность. Затем мочевина большей частью вновь переходит в нисходящую и восходящую тонкие части петли Генле (медулло-медуллярная рециркуляция) и частично реабсорбируется в прямых сосудах
Реабсорбция глюкозы В почке D-глюкоза практически полностью реабсорбируется (рис. 9-13). Местом реабсорбции является проксимальный каналец. В настоящее время описана молекулярная структура ответственных за это белков-переносчиков. В апикальной мембране проксимального извитого канальца в сегменте S1 работает механизм вторичного активного транспорта, который осуществляется белком-переносчиком SGLT2 (sodium-glucose transporter, Typ2: SGLT2), обладающем низкой аффинностью. Он одновременно переносит Na+ и глюкозу (но не галактозу) в соотношении 1:1. С помощью данного переносчика к концу проксимального извитого канальца (например, у крысы) реабсорбируется около 95% профильтровавшегося количества глюкозы. В сегменте S3 прямой части проксимального канальца обнаружен еще один белок-переносчик SGLT1, расположенный на апикальной мембране, который обладает высокой аффинностью и на одну молекулу глюкозы переносит два иона Na+ (котранспорт). С помощью этого переносчика концентрация глюкозы в просвете канальца может быть снижена настолько, что в конечной моче обнаруживается лишь 1/1000 профильтровавшегося количества глюкозы. Переход глюкозы, поступившей из просвета канальца, в кровь околоканальцевых капилляров является пассивным. Он обеспечивается (независимым от ионов) переносчиком GLUT2 (glucose transporter 2: GLUT2) и движим химическим градиентом глюкозы (так называемая облегченная диффузия). GLUT2 также может переносить галактозу и фруктозу, при этом галактоза переносится в клетку из просвета канальца в результате вторично-активного транспорта с помощью SGLT1, а фруктоза - пассивно с помощью переносчика GLUT5. Рис. 9-13. Транспорт глюкозы. Реабсорбция глюкозы происходит большей частью в начальных отделах проксимального канальца. Если увеличивается профильтровавшееся количество глюкозы, то по мере насыщения переносчиков в начальных отделах проксимального канальца в процесс реабсорбции вовлекаются дистальные сегменты проксимального канальца, и ценная для организма глюкоза практически полностью реабсорбируется
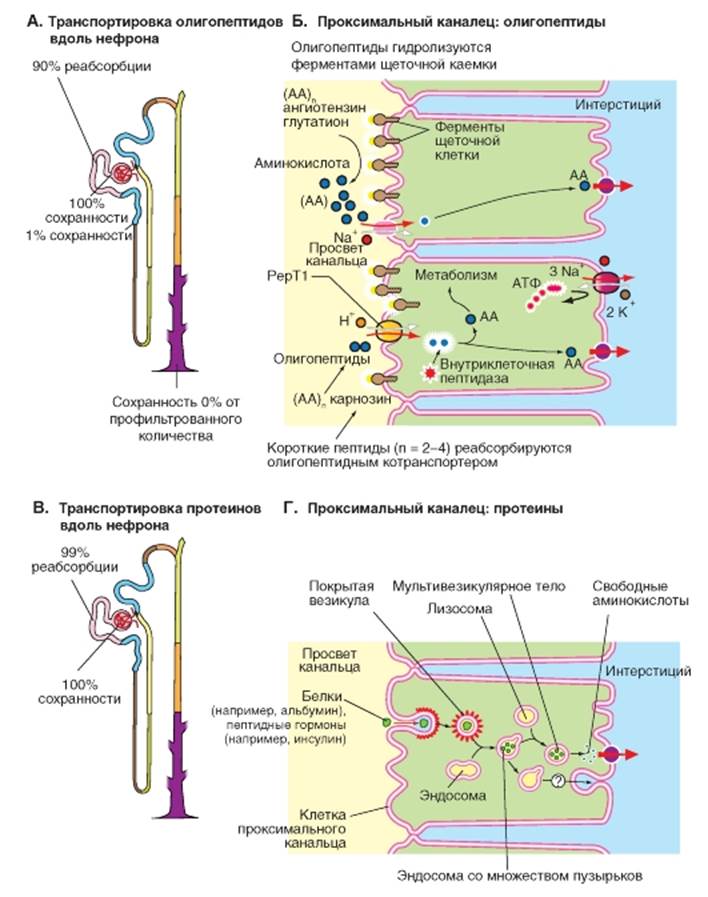
Реабсорбция аминокислот Аминокислоты реабсорбируются на >98% (отдельные аминокислоты, например, L-валин до >99,8%). Исключениями являются глицин (96%), гистидин (94%), а также таурин (около 90%). Для аминокислот существует множество белков-переносчиков, которые обладают специфичностью к одной группе структурно-родственных L-аминокислот (рис. 9-14). Так в апикальной мембране клеток эпителия проксимального канальца имеются белки-переносчики, обеспечивающие сопряженный перенос Na+ и аминокислот (котранспорт): а) для анионных аминокислот, таких как L-глю- тамат- и L-аспартат- (переносит 2Na+/анион аминокислоты-); б) для большинства нейтральных L-аминокис- лот (1Na+/нейтральная аминокислота; высокая эффективность); в) для L-пролина; г) для β-аминокислот, таких как таурин, β-аланин и др. Благодаря им внутриклеточные концентрации аминокислот увеличиваются в несколько раз по сравнению с концентрациями в плазме крови: таурина, например, - в 30 раз, а L-глутамата- - в 50 раз. Поэтому выход аминокислот из клетки в интерстициальное пространство и далее к капиллярам может осуществляться пассивно («облегченная диффузия» за счет различных переносчиков). Катионные аминокислоты L-аргинин+, L-ли- зин+ и L-орнитин+ могут проникать в клетку как пассивно (мембранный потенциал как движущая сила!), так и переноситься в результате вторично-активного транспорта совместно с Na+. Такой переносчик (называемый D2H) связывает и переносит также цистеин и другие нейтральные аминокислоты. Как, вопреки направленному против них потенциалу, катионные аминокислоты покидают клетку через базолатеральную мембрану, пока неясно. Переносчики аминокислот стереоспецифичны, хотя есть и исключения (например, транспорт D-аспартата переносчиком анионных аминокислот). Как переносчик глюкозы принимает лишь D- (но не L-глюкозу), так и в случае аминокислот транспортируются лишь L- (но не D-) изомеры. Поскольку сходные аминокислоты переносятся одним и тем же переносчиком, одна аминокислота (например, аргинин+) может тормозить реабсорбцию другой аминокислоты (в данном случае, лизина+). Рис. 9-14. Транспорт аминокислот. Реабсорбция аминокислот происходит большей частью в начальных отделах проксимального канальца. В начальных отделах проксимального канальца в процесс реабсорбции вовлекаются дистальные сегменты проксимального канальца, и ценные для организма аминокислоты практически полностью реабсорбируются. В качестве примера на рисунке представлены лишь отдельные переносчики. В общем случае переносчик для аминокислот обозначен как АА - аминокислоты. Далее представлены отдельные примеры переносчиков: Glu - для глутамата, Lys - для лизина, Pro - для пролина
Реабсорбция олигопептидов и белков Пептиды реабсорбируются расщепленными и нерасщепленными. Некоторые ди- и трипептиды (например, карнозин) устойчивы по отношению к действию пептидаз просвета канальца. Для них в апикальной мембране клеток проксимального канальца имеется два переносчика, обеспечивающих перенос пептида и H+ в клетку (котранспорт): PерT1 (сегмент S1) и PерT2 (сегмент S2). С помощью этих переносчиков дипептиды (и определенные антибиотики - цефалоспорины) могут в результате вторичного активного транспорта переноситься через щеточную каемку в клетку по направлению градиента H+ (рис. 9-15 Б). Такие пептиды в большинстве случаев расщепляются до аминокислот внутриклеточно. Для коротких пептидных цепочек существует еще одна форма реабсорбции. В щеточной каемке проксимального канальца одновременно с рядом ферментов (например, мальтаза, трегалаза) обнаруживается высокая активность аминопептидаз, эндопептидаз и γ-глютамилтрансфераз (γ-GT), которые действуют в просвете нефрона. Они способны настолько быстро расщеплять белки в просвете канальца, что при прохождении мочи через проксимальный каналец (около 12 с) остается еще достаточно времени для реабсорбции образующихся продуктов расщепления, т.е. аминокислот. Для больших белков клубочковый фильтр не-проницаем, особенно если они заряжены отрицательно. Концентрации в фильтрате количественно важного белка плазмы крови альбумина составляет лишь 0,01-0,05% от его концентрации в плазме крови (около 40 г/л). Несмотря на слабую проницаемость фильтра, профильтровавшееся количество альбумина при GFR 180 л/ сут может достигать (180 x 40 x 0,0001 соответственно = 0,0005) 0,75-4 г/сут. К нему могут быть добавлены другие белки плазмы, в особенности низкомолекулярные: лизоцим, обломки иммунглобулинов, α1- и β2-микроглобулин и т.д. В моче обычно появляется лишь 35 мг альбумина в день. Это означает, что более чем 96% профильтровавшегося альбумина реабсорбируется в проксимальном канальце (подобное справедливо и для других белков). В то время как короткие пептидные цепочки гидролизуются уже в просвете канальца, большие пептиды с дисульфидными мостиками (такие как инсулин, β2-микроглобулин) и белки, такие как альбумин, могут реабсорбироваться в проксимальном канальце за счет эндоцитоза, который запускается взаимодействием белка со специфическим рецепторным комплексом щеточной каемки (рис. 9-15 Г). Этот процесс идет с потреблением АТФ. Белки связываются с рецепторами (=комплекс мегалинкубилин) щеточной каемки и транспортируются к основанию микроворсинок, где отшнуровываются эндоцитозные везикулы, превращающиеся внутри клетки в эндосомы. Эндосомы сливаются с лизосомами, и протеазы лизосом расщепляют захваченные белки до аминокислот. (Некоторые белки гидролизируются уже в эндосомах). Появляющиеся при этом аминокислоты посредством переносчика везикулярной мембраны переносятся в цитоплазму. Мембрана везикулы, несущая рецепторы, снова встраивается в плазматическую мембрану, обращенную в просвет канальца (мембраный цикл). С помощью опосредованного мегалин-кубилином эндоцитоза реабсорбируются также некоторые связанные с белками витамины, например ретинол (на ретинол-связывающем белке), кобаламин (на транс-кобаламине) и 25-OH-холекальциферол (=кальцидол; на белке, связывающем витамин D, DBP). Так 25-OH-холекальциферол попадает в клетки, и под действием 1-α-гидроксилазы превращается в кальцитриол. Рис. 9-15. Транспорт олигопептидов и белков. Рассматривается канальцевая реабсорбция олигопептидов и белков. Большинство коротких пептидных цепочек (например, глюкагон, ангиотензин II, рилизинг факторы и глутатион) настолько быстро гидролизуются пептидазами щеточной каемки, действующими в просвете канальца, что образующиеся при этом аминокислоты могут быть реабсорбированы, прежде чем они достигнут конца проксимального канальца. Определенные ди- и трипептиды (например, карнозин) более устойчивы к действию пептидаз. Поэтому в проксимальном канальце они транспортируются внутрь клетки белком-переносчиком, осуществляющим сопряженный перенос пептид-H+ (котранспорт), где и расщепляются (А). Белки, такие как лизоцим, β 2-микроглобулин и альбумин, а также пептиды, содержащие дисульфидные мостики (например, инсулин), реабсорбируются в проксимальном канальце посредством эндоцитоза, который запускается взаимодействием белка со специальными рецепторами, и гидролизуются в лизосомах
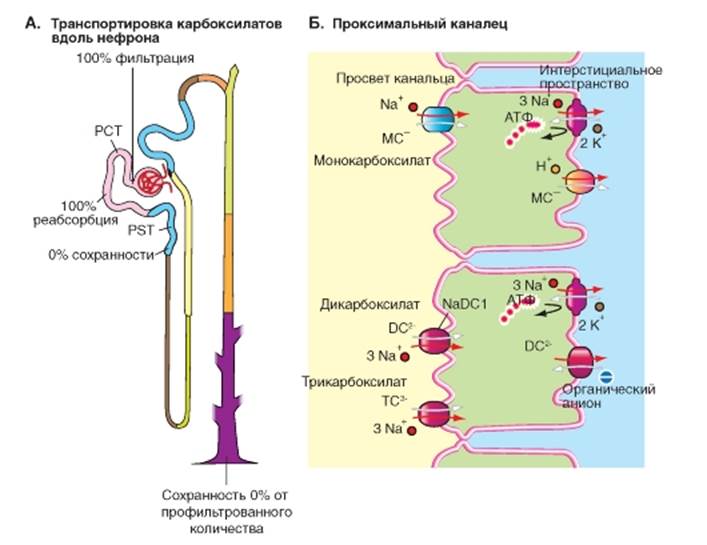
Транспорт трикарбоксилатов Дикарбоксилаты появляются в клетке либо в результате обмена веществ, протекающего в клетках проксимального канальца, или переносятся в клетку из внеклеточного пространства в результате вторично-активного транспорта с помощью переносчика hNaDCl, который осуществляет сопряженный перенос Na+ - дикарбоксилат (котранспорт, рис. 9-16). В последнем случае перенос OA- представляет собой «третично»-активный транспорт. Для секреции амфифильных конъюгатов (например, сцепленных с глутатионом липофильных токсинов) в апикальной мембране, отделяющей клетку от просвета канальца, дополнительно существует АТР-зависимый конъюгатный насос MRP2 (Multi-drug-resistance protein, Typ 2: MRP2). Рис. 9-16. Транспорт моно-, ди- и трикарбоксилатов
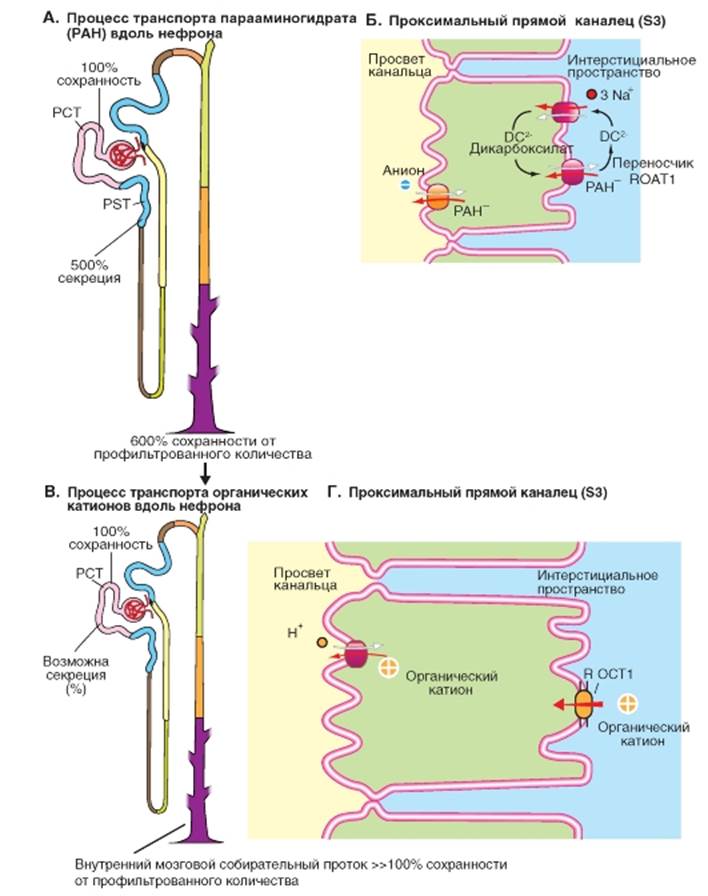
Секреция органических ионов а из клетки выносит дикарбоксилаты (2 оксиглутарат2-, сукцинат2-; антипорт). На апикальной мембране имеется обменник, который выводит из просвета канальца анионы (например, ураты) в обмен на РАН, который секретируется в просвет канальца. Секреция органических катионов, OK+ (органические основания) также осуществляется в проксимальном канальце (рис. 9-17 В, Г). К органическим катионам относится ряд растительных алкалоидов (таких как атропин и морфин), диуретик амилорид или присущий организму гистамин. Рассмотрим пример, представленный на рис. 9-17 В, Г. В этом случае механизм вторичного активного транспорта локализован в апикальной мембране клеток эпителия канальца, где путем переноса обмениваются органические катионы на Н+(противоположно направленный транспорт или антипорт). Движущей силой этого процесса является электрохимический градиент Н+через апикальную мембрану со стороны просвета канальца, который поддерживается переносчиком, осуществляющим обмен Na+ на Н+(антипорт) и Н+-АТФазой. Дополнительно, по всей видимости, возможна первично-активная секреция OK+ посредством MDR1 (Multi Drug Resistance Protein Type 1: MDR1). Переход OK+ из интерстициального пространства в клетку через базолатеральную мембрану осуществляется за счет полиспецифичного переносчика органических катионов ROCT1 (renal organic cation transporter Typ1: ROCT1; облегченная диффузия). Рис. 9-17. Секреция органических анионов и катионов. A, Б - органические анионы (OA-, в качестве примера на рисунке продемонстрирована PAH-) и B, Г - органические катионы (OK+) попадают в просвет проксимального канальца не только в результате клубочковой фильтрации, но и в результате активной секреции. Поскольку OA- и OK+ практически не реабсорбируются, то они могут быть выведены с мочой в больших количествах Если говорить коротко, то в мембране клеток проксимального канальца имеются переносчики, которые активно секретируют в просвет канальца органические кислоты и основания. Таким образом, разнообразные конечные продукты обмена веществ, чужеродные и ядовитые вещества могут быть быстро выведены из организма. Интенсивность процессов секреции может быть настолько высокой, что выводимое количество оказывается в четыре раза выше профильтрованного. Итак, к задачам проксимального канальца относится также секреция органических веществ. Эта секреция может очень сильно ускорять процесс выведения веществ (по сравнению с только фильтрующимися веществами), поскольку к профильтровавшемуся количеству вещества добавляется секретируемое количество вещества. Секреция органических анионов, ОА- (органические кислоты), таких как парааминогиппуровая кислота (РАН), мочевая кислота, гиппуровая кислота, пенициллин, фуросемид, индометацин и различных конъюгатов токсинов, осуществляется путем вторичного активного транспорта. Рассмотрим пример для РАН, представленный на рис. 9-17 А, Б. В базолатеральной мембране имеется по крайней мере один переносчик органических анионов ROAT1 (ROAT1, renal organic anion transporter Typ1). ROAT1 связываясь с ОА-переносит его через базолатеральную мембрану в клетку,
Транспорт уратов Ураты - кислые, хорошо растворимые натриевая и калиевая соли мочевой кислоты. Высокая растворимость уратов в плазме объясняется наличием связывающих белков и, возможно, некоторых растворяющих низкомолекулярных соединений. По мере продвижения по системе почечных канальцев часть экскретируемых с мочой уратов превращается в мочевую кислоту: 2,6,8-триоксипу- рин, или C5H4N4O3. Мочевая кислота является конечным продуктом пуринового обмена, причем ксантиноксидаза катализирует два последних этапа образования мочевой кислоты (гипоксантин - ксантин - урат). В проксимальном канальце мочевая кислота одновременно реабсорбируется и секретируется; процессы реабсорбции превалируют, поэтому около 10% профильтровавшегося количества мочевой кислоты выводится с мочой. При высококонцентрированной моче это означает, что концентрация мочевой кислоты в 20-30 раз превышает ее концентрацию в плазме крови, нормальное значение для которой составляет около 0,25 ммоль. Повышение концентрации мочевой кислоты в плазме крови (гиперурикемия, концентрация от 0,4 и выше 0,6 ммоль) может быть вызвано несколькими причинами: уменьшением выделения почками (ослабленная секреция или усиленная реабсорбция); образованием больших количеств мочевой кислоты в результате обмена веществ, например при богатом пуринами питании (мясо, внутренности); определенными дефектами ферментов; болезнями, вызывающими многочисленную гибель клеток. Мочевая кислота и ее соли при низком значении рН плохо растворимые. Поэтому при гиперурикемии они выпадают в виде кристаллов, следствием чего является тяжелое поражение органов. На рис. 9-18 представлена и реабсорбция и секреция уратов, при этом реабсорбция обычно доминирует. Прописанный на рисунке путь (1) включает в себя обменник уратов на ОН- или НСО3- со стороны просвета проксимального канальца, расположенный на апикальной мембране (третично активный транспорт - tertiary active transport) и работающий параллельно с апикальным Na-H-обменником (вторично активный транспорт). Второй путь (2) представляет собой обменник уратов на монокарбоксилат (лактат, β -гидроксибутират) или дикарбоксилат (третично активный транспорт) в параллель с работающим по Na+ карбоксилатным контранспортом (вторично активный транспорт - разница градиента Na+). Третий путь (3) - обменник уратов со стороны просвета проксимального канальца на анионы, такие как РАН (третично активный транспорт), и обменник на базолатеральной мембране, осуществляющий усвоение РАН-подобных анионов (вторично активный транспорт) в обмен на соли мочевой кислоты. Рис. 9-18. Транспорт уратов (солей мочевой кислоты).
|
|||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-20; просмотров: 814; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.137 (0.021 с.) |