Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Стрелки указывают направления изменений от контрольных значений потенциалов действия и амплитуды силы сокращенияСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Влияние нейтротрансмиттеров, их агонистов и антагонистов на клетки синоатриального узла Нейротрансмиттеры вегетативной нервной системы воздействуют на автоматию, изменяя клеточные трансмембранные ионные токи. Адренергические трансмиттеры увеличивают все три тока, участвующих в автоматии SA-узла. Чтобы увеличить крутизну диастолической деполяризации, усиление If и ICa адренергическими трансмиттерами должно превысить усиление IK. Адренергические трансмиттеры могут существенно влиять на GCa. Так, адренергический нейротрансмиттер норадреналин или, например, агонист β-адренергического рецептора изопротеренол (рис. 6-13 А) и различные другие катехоламины могут усиливать Ca2+-проводимость. Увеличение Ca2+-проводимости катехоламинами, возможно, является основным механизмом, по которому они усиливают сократимость сердечной мышцы. Гиперполяризация (рис. 6-13 Б), вызванная ацетилхолином, который высвобождается из окончаний блуждающего нерва, достигается увеличением GK. Это изменение в проводимости опосредуется через активацию специфических K+-каналов, регулируемых ацетилхолином. Эти каналы регулируются под действием ацетилхолина (например, внеклеточной концентрацией ацетилхолина). В сердце имеются мускариновые K+-каналы (KACh) - тип KG каналов, активирующийся G-белками, ответственными за эффект ацетилхолина. Возникающий ток обозначают как IKACh. В сердце ацетилхолин также понижает If и ICa токи. У здоровых людей, находящихся в спокойном состоянии, преобладает обычно тоническое влияние парасимпатической нервной системы. Когда здоровому человеку вводят атропин, антагонист мускариновых рецепторов, который блокирует воздействия парасимпатической нервной системы, частота сердечных сокращений обычно значительно возрастает. Если здоровому человеку вводят пропранолол, антагонист β-адренергических рецепторов, который блокирует воздействия симпатической нервной системы, частота сердечных сокращений незначительно уменьшается. Когда оба отдела вегетативной нервной системы блокированы, частота сердечных сокращений у молодых людей в среднем достигает значения 100 ударов в минуту. Частота сердечных сокращений, которая устанавливается при полной блокаде вегетативной нервной системы, называется собственной частотой сердечных сокращений. Рис. 6-13. Изменение ритма пейсмейкерной активности изолированной синоатриальной клетки под действием низких концентраций агониста β -адренорецепторов изопротеренола (ISO) (А) и агониста мускариновых холинорецепторов ацетилхолина (ACh) (Б). Ускорение или замедление ритма происходят, главным образом, за счет активации изопротеренолом или подавлением ацетилхолином пейсмейкерного тока If
Механизм влияния норадреналина и ацетилхолина Различные факторы, такие как нейротрансмиттеры и лекарственные средства, могут существенно влиять на проводимость каналов. Чтобы усилить Ca2+ проводимость, катехоламины сначала взаимодействуют с β -адренергическими рецепторами мембраны клетки сердца. Это взаимодействие стимулирует мембранно-связанный фермент аденилилатциклазу, что повышает внутриклеточную концентрацию циклического аденозин монофосфата (цАМФ). Повышение уровня цАМФ усиливает активацию Ca2+-каналов L-типа в клеточной мембране и таким образом увеличивает вход Ca2+ в клетки из межклеточной жидкости. Ацетилхолин взаимодействует с мускариновыми рецепторами клеточной мембраны, чтобы наоборот ингибировать аденилилатциклазу. Таким путем ацетилхолин противодействует активации Ca2+-каналов, уменьшая, таким образом, GCa. Синоатриальный и атриовентрикулярный узлы содержат много холиностеразы, фермента, разрушающего ацетилхолин. Ацетилхолин, высвобож- денный из окончаний блуждающих нервов быстро гидролизируется. Благодаря быстрому разрушению ацетилхолина воздействия, вызываемые любой стимуляцией блуждающего нерва, очень быстро прекращаются после окончания стимуляции. Кроме того, влияние блуждающего нерва на деятельность синоатриального или атриовентрикулярного узлов имеет очень короткий латентный период (от 50 до 100 мс), потому что выделяемый ацетилхолин активирует специфические ацетилхолин-регулируемые К+-каналы (IKACh) в клетках сердца. Эти каналы открываются так быстро потому, что ацетилхолин действует минуя систему вторичных мессенджеров такую, как система аденилатциклазы. Сочетание двух характерных особенностей блуждающих нервов - короткого латентного периода и быстрого угасания ответной реакции - позволяет блуждающим нервам регулировать деятельность синоатриального и атриовентрикулярного узлов при каждом сокращении сердца. В области синоатриального узла влияние парасимпатической нервной системы обычно превосходит влияние симпатической нервной системы. Рис. 6-14. Влияние агонистов α1- и β -адренорецепторов и М2-холинорецепторов через вторичные внутриклеточные посредники на потенциал-зависимые ионные токи IK, If и ICa,L сердечных клеток. Кроме того, стимуляция М2-холинорецептора приводит к активации выходящего калиевого тока IKACh. АЦ - аденилатциклаза, ФДЭ - фосфодиэстераза, СПР - саркоплазматический ретикулум, IP3 - инозитолтрифосфат, DАG - диацилглицерол, PKA и PKC - протеинкиназы A и C, соответственно
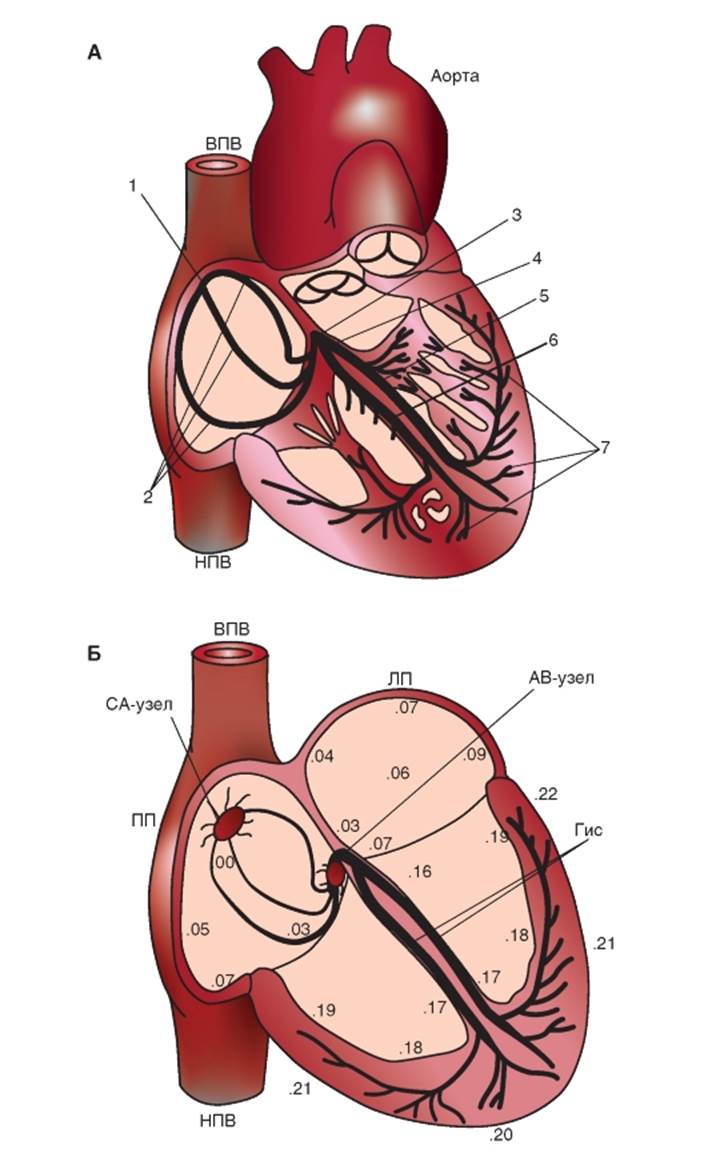
Структура проводящей системы сердца Автоматия - это способность к самовозбуждению, или способность генерировать свое собственное возбуждение и сокращение. Регулярность такой пейсмейкерной активности определяет ритмичность работы сердца. Доказательством наличия автоматизма служит сокращение сердца, полностью извлеченного из организма. Областью сердца млекопитающих, которая обычно генерирует импульсы с самой большой частотой, является синоатриальный (SA) узел, это главный водитель ритма сердца. Детализированное картирование электрических потенциалов на поверхности правого предсердия показывает, что существуют две или три области автоматии, находящиеся в 1 или 2 см от SA-узла, и составляют совместно с SA-узлом предсердный пейсмейкерный комплекс. Иногда все эти локусы одновременно инициируют импульсы. А иногда, область самого раннего возбуждения сдвигается от локуса к локусу, в зависимости от определенных условий, таких как уровень активности автономной нервной системы. Из SA-узла сердца импульсы радиально распространяются по всему правому предсердию по обычным предсердным волокнам миокарда со скоростью проведения около 1 м/с. Специальный путь, передний межпредсердный пучок миокарда (или пучок Бахмана), проводит импульс из SA-узла непосредственно к левому предсердию. Волна возбуждения, которая направляется вниз через правое предсердие, в конечном счете, достигает атриовентрикулярного (AV) узла, который является обычно единственным путем прохождения сердечного импульса к желудочкам. Волна возбуждения из предсердий достигает желудочков через AV-узел. У взрослых людей этот узел около 22 мм в длину, 10 мм в ширину и 3 мм в толщину. Узел расположен сзади, на правой стороне межпредсердной перегородки, вблизи устья коронарного синуса. AV-узел содержит те же самые два типа клеток, что и SA-узел, но круглые клетки в AV-узле менее распространены, а преобладают удлиненные. AV-узел состоит из трех функциональных областей: (1) область AN (atrium-nodus), переходная зона между предсердием и остальной частью узла; (2) N (nodus) средняя часть AV-узла; и (3) NH (nodus-His) - зона, в которой волокна узла постепенно сливаются с пучком Гиса, и которая представляет собой верхнюю часть специализированной проводящей системы желудочков. Обычно AV-узел и пучок Гиса являются единственными проводящими путями, по которым импульсы сердца проходят от предсердий к желудочкам. Пучок Гиса проходит субэндокардиально вниз, приблизительно на 1 см, по правой стороне межжелудочковой перегородки и затем разделяется на правую и левую ножки пучка. Правая ножка пучка, которая является прямым продолжением пучка Гиса, направляется вниз по правой стороне межжелудочковой перегородки. Левая ножка пучка, которая значительно толще, чем правая, отходит почти перпендикулярно от пучка Гиса и проникает через межжелудочковую перегородку. На субэндокардиальной поверхности левой стороны межжелудочковой перегородки, левая ножка пучка разделяется на тонкую переднюю и толстую заднюю ветви. Правая ножка пучка и две ветви левой ножки пучка в конечном счете ветвятся, образуя сложную сеть проводящих волокон, названных волокнами Пуркинье, которые распространяются по субэндокардиальным поверхностям обоих желудочков. У некоторых видов млекопитающих, например у крупного рогатого скота, сеть волокон Пуркинье организована в виде дискретных, инкапсулированных волокон. Волокна Пуркинье имеют множество линейно организованных саркомеров, таких же, как у кардиомиоцитов. Однако T-тубулярная система отсутствует в волокнах Пуркинье у многих видов, хотя она хорошо развита в кардиомиоцитах. Волокна Пуркинье - самые крупные клетки сердца: они имеют диаметр от 70 до 80 мкм, в то время как диаметр кардиомиоцитов желудочков лежит в диапазоне от 10 до 15 мкм. Частично из-за своего большого диаметра, скорость проведения возбуждения (от 1 до 4 м/с) в волокнах Пуркинье превышает таковую у любого другого типа сердечных волокон. Более высокая скорость проведения обеспечивает быструю активацию всей эндокардиальной поверхности желудочков. Рис. 6-15. Проводящая система сердца. А - строение проводящей системы сердца: 1 - синоатриальный узел, 2 - проводящие пути предсердий, 3 - атриовентрикулярный узел, 4 - пучок Гиса, 5 и 6 - правая и левая ножки пучка Гиса, 7 - волокна Пуркинье, ВПВ и НПВ - верхняя и нижняя полые вены. Б - скорость распространения возбуждения по миокарду предсердий и желудочков. Цифры указывают время (в сундах) от момента возникновения возбуждения в синоатриальном узле до начала возбуждения в данном отделе миокарда. Скорость проведения возбуждения в синоатриальном узле - 1-2 см/с, в предсердиях около 1 м/с, в атриовентрикулярном узле - 1-5 см/с, в проводящей системе Гиса-Пуркинье - 2-4 м/с. Обозначения: СА - синоатриальный узел, АВ - атриовентрикулярный узел, Гис - ножки пучка Гиса, ВПВ и НПВ - верхняя и нижняя полые вены, ПП и ЛП - правое и левое предсердия
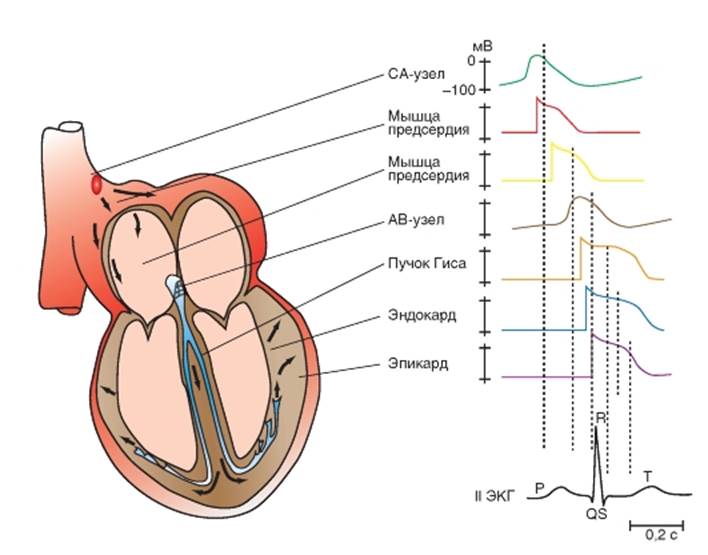
Функции и свойства проводящей системы сердца Проводящая система сердца обеспечивает генерацию ритма возбуждений, последовательность и синхронность возбуждения, а потом и сокращения. Все это повышает эффективность работы сердца как насоса. Разные отделы проводящей системы выполняют различную функцию, что определяет их свойства. Синоатриальный узел ответственен за генерацию потенциалов и в норме является водителем ритма. Основным свойством атриовентрикулярного узла является проведение возбуждения. Возбуждение в АV-узле пройдет только в том случае, если оно произойдет сразу в нескольких клетках. Необходимо еще обратить внимание на важное свойство АV-узла: это значительное снижение скорости проведения возбуждения до 0,02-0,05 м/с. Это снижение скорости происходит на участке длиной 1,5-2 мм и обусловлено особенностями распространения потенциала с медленным ответом. Необходимость значительного снижения скорости очевидна, поскольку механические процессы в сердце протекают значительно медленнее, чем электрические, и нужно время, чтобы кровь успела из предсердий попасть в желудочки. Наименьшая скорость проведения имеет места в средней части (nodus). АV-узел обладает односторонней проводимостью. Атриовентрикулярный узел помимо проведения возбуждения при необходимости может взять на себя функцию водителя ритма. Кроме того, особые свойства АУ-узла обеспечивают неполную блокаду при проведении возбуждения свыше 180-200 в минуту. Далее возбуждение идет по пучку Гиса и его ножкам со скоростью 2-3 м/с, переходит на волокна Пуркинье, здесь скорость максимальная - 4-5 м/с и, наконец, достигает клеток рабочего миокарда. Скорость распространения возбуждения от субэндокардиальных окончаний волокон Пуркинье по рабочему миокарду желудочков составляет около 1 м/с. На рис. 6-16 продемонстрированы различные типы потенциалов действия, зарегистрированные последовательно от различных участков сердца и сопоставлены с одновременной записью электрокардиограммы. На данном рисунке отчетливо видно, что наибольшая задержка проведения возбуждения имеет место на участке: предсердие - атриовентрикулярный узел. Рис. 6-16. Последовательная регистрация потенциалов действия от различных участков сердечной мышцы и сопоставление их с электрокардиограммой, зарегистрированной во втором стандартном отведении
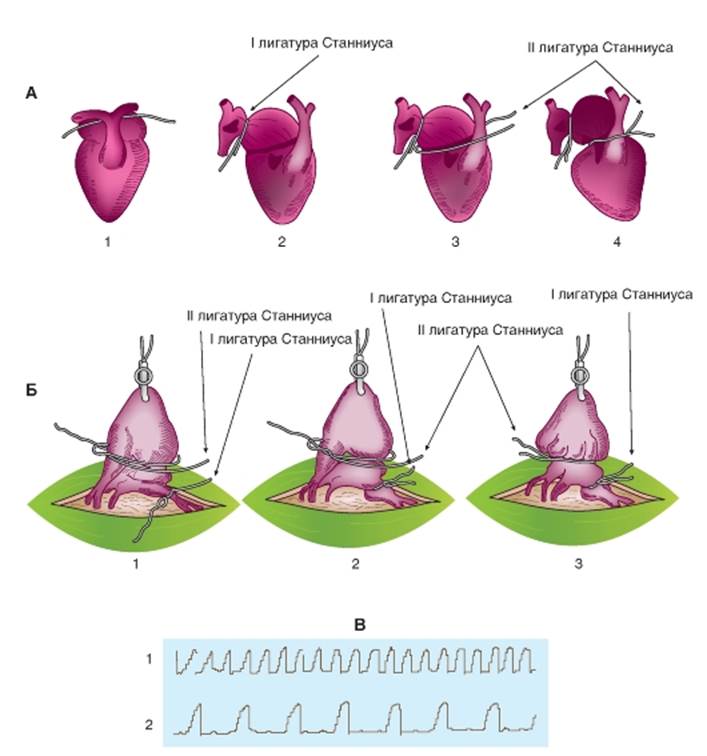
Автоматия различных отделов сердца Проводящая система холоднокровных животных включает два узла автоматии: синусный, расположенный в венозном синусе, и атриовентрикулярный - на границе предсердий и желудочка. Все участки проводящей системы обладают способностью к автоматии, которая изменяется от основания сердца к верхушке (градиент автоматии). Наибольшей автоматией обладает синоатриальный узел (синусный - у холоднокровных). Его называют центром автоматии первого порядка. При нарушении функции синоатриального узла какими-либо факторами проявляется автоматия атриовентрикулярного узла. Ритм сокращений сердца, задаваемый этим узлом, вдвое и более раз реже, чем в норме. После выключения атриовентрикулярного узла роль водителя ритма переходит к пучку Гиса, сокращения сердца становятся еще реже и т.д. Автоматия вторичных узлов проявляется не сразу после выключения узла высшего порядка, а через промежуток времени, который длится от нескольких сунд до нескольких минут, - это так называемая преавтоматическая пауза. Для изучения проводящей системы сердца лягушки используют метод наложения лигатур на области расположения центров автоматии (лигатуры Станниуса). Применяют две лигатуры Станниуса (рис. 6-17 A). Первая лигатура накладывается в виде тугой перевязки между венозным синусом и предсердиями. Она отделяет синусный узел от остальных отделов сердца и препятствует доступу возбуждения к ним. Вследствие этого сокращения предсердий и желудочка прекращаются. Венозный синус продолжает сокращаться в исходном ритме. Опыт доказывает наличие в венозном синусе ведущего узла автоматии. Вторая лигатура Станниуса накладывается между предсердиями и желудочком, она служит для раздражения атриовентрикулярного узла и пробуждения его автоматии. После наложения второй лигатуры возникают сокращения предсердий и желудочка, но в ином ритме, чем сокращения синуса. Опыт доказывает существование второго узла автоматии, расположенного в атриовентрикулярной области. Регистрируют работу сердца в норме с подведенными лигатурами Станниуса. Затем туго завязывают лигатуру на границе между предсердиями и венозным синусом (рис. 6-17 Б). Сразу после перетяжки предсердия и желудочек останавливаются, а сокращения венозного синуса продолжаются в прежнем ритме. Результаты наложения первой лигатуры регистрируют на самописце. Слабые сокращения венозного синуса можно записать только при очень хорошо отпрепарированном препарате. На некоторых препаратах удается иногда обнаружить самостоятельное проявление автоматии атриовентрикулярного узла через какое-то время после наложения первой лигатуры Станниуса (иногда через несколько минут). Ритм спонтанно возникающих сокращений меньше исходного синусного ритма. Если же автоматия атриовентрикулярного узла спонтанно не восстанавливается, можно ускорить ее проявление каким-либо внешним стимулом. Таким раздражителем может служить вторая лигатура Станниуса, которая накладывается в области атриовентрикулярной борозды (рис. 6-17 Б). Вторую лигатуру затягивают нетуго, до появления сокращений сердца. Вначале эти сокращения, как правило, довольно часты, но постепенно урежаются, и вскоре устанавливается истинный атриовентрикулярный ритм, который в 2-3 раза реже исходного ритма сердца (рис. 6-17 В2). Рис. 6-17. Изучение автоматии различных отделов сердца лягушки - опыт Станниуса. А - выключение отдельных узлов проводящей системы с помощью лигатур Станниуса: 1 - подведение лигатуры для перевязки венозного синуса; 2 - венозный синус отделен лигатурой от предсердий; 3 - подведена вторая лигатура для отделения предсердий от желудочков; 4 - предсердия отделены от желудочков. Б - схема наложения лигатур Станниуса на сердце лягушки. Принцип наложения лигатур Станниуса с целью получения искусственного блока сердца (I - первая лигатура Станниуса; II - вторая лигатура Станниуса): 1 - подведение первой и второй лигатур Станниуса, 2 - затягивание первой лигатуры Станниуса, 3 - затягивание второй лигатуры Станниуса. В - механограмма сердца лягушки: 1 - механограмма сердца лягушки в норме, 2 - механограмма после искусственного поперечного блока Сердца
Влияние температуры на частоту сокращений сердца Доказательством автоматии сердца служит тот факт, что вырезанное из организма сердце продолжает ритмически сокращаться. Сердце имеет несколько центров автоматизма, однако используя факт влияния температуры на скорость течения физиологических процессов, можно доказать ведущую роль синусного узла в автоматизме сердца. Если нагревать или охлаждать различные отделы сердца лягушки, то выявляется изменение частоты его сокращений только при изменении температуры синуса, тогда как изменение температуры других отделов сердца (предсердий, желудочка) сказывается лишь на силе мышечных сокращений. Эффект влияния температуры можно наблюдать и на изолированном, и на неизолированном сердце лягушки. У обездвиженной лягушки обнажают сердце (рис. 6-18 А) собирают установку по схеме, представленной на рис. 6-18 Б. Захватывают серфином или лигатурой верхушку сердца и устанавливают штырь регистратора в горизонтальном положении, опуская или поднимая дощечку с лягушкой. К основанию сердца, находящегося в вертикальном положении (к зоне синусного узла), подводят термод, заполненный водой комнатной температуры. С помощью механоэлектрического преобра- зователя записывают на самописце кривую сокращений сердца - механограмму (рис. 6-18 В1). По термоду пропускают воду температурой 4 °С и продолжают запись механограммы, регистрируя изменение механограммы сердца (рис. 6-18 В2). После получения устойчивого эффекта вновь пропускают через термод воду комнатной температуры для получения исходной механограммы (рис. 6-18 В3). Затем пропускают через термод воду температурой около 36 °С и продолжают запись механограммы, регистрируя изменение механограммы сердца (рис. 6-18 В4). После получения эффекта пропускают через термод воду комнатной температуры для получения исходной механограммы (рис. 6-18 В5). Аналогичным образом проводят исследование влияния температуры на частоту сокращений сердца, перемещая термод к другим участкам сердца (желудочку). После прекращения температурного воздействия изолируют сердце из организма, для чего предварительно перевязывают сосуды и перерезают их ниже места перевязки. Перерезают полую вену, следя за тем, чтобы не повредить венозный синус. Если изоляция сделана правильно, то сердце, вырезанное из тела лягушки и помещенное в физиологический раствор, будет сокращаться продолжительное время. Рис. 6-18. Влияние нагревания и охлаждения венозного синуса на частоту сокращений сердца лягушки (опыт Гаскелла).
|
||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-20; просмотров: 509; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.219.127.59 (0.009 с.) |