Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
На рисунке изображены большой (красным) и малый (синим) круги кровообращения с параллельным и последовательным расположением кровеносных сосудов. Капилляры изображены тонкими линиямиСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
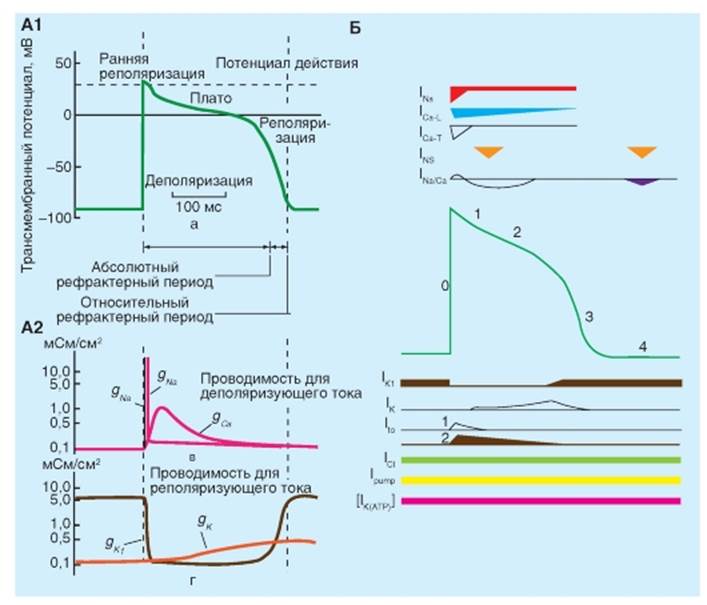
Типы и характеристика потенциалов действия кардиомиоцитов Изменения потенциала, отводимого от типичного мышечного волокна желудочка, показаны на рис. 6-3 А. Когда два электрода помещены в раствор электролита рядом с полоской покоящейся сердечной мышцы, то между этими двумя электродами не регистрируется разность потенциалов (точка а). В точке b, когда один из электродов введен внутрь мышечного волокна сердца, измерительный прибор мгновенно регистрирует разность потенциалов (Vm) между внутренней и внешней стороной клеточной мембраны. Потенциал внутри клетки примерно на -90 мВ ниже, чем у окружающей среды. В точке c клетка желудочка возбуждается электрическим стимулом, и клеточная мембрана быстро деполяризуется. Во время деполяризации разность потенциалов фактически меняет знак таким образом, что потенциал внутри клетки превышает таковой снаружи приблизительно на 20 мВ. Быстрое нарастание потенциала действия обозначается как фаза 0. За нарастанием мгновенно следует короткий период частичной ранней реполяризации (фаза 1) и затем плато (фаза 2), которое удерживается приблизительно от 0,1 до 0,2 с. Мембрана затем реполяризуется (фаза 3) до тех пор, пока поляризация снова не достигнет (в точке e) состояния покоя (фаза 4). Окончательная реполяризация (фаза 3) развивается более медленно, чем деполяризация (фаза 0). В сердце регистрируют два основных типа по-тенциалов действия, которые показаны на рис. 6-3. Один тип, быстрый ответ (рис. 6-3 А), возникает в рабочих миоцитах предсердий и желудочков и в специализированных проводящих волокнах (волокна Пуркинье сердца). Другой тип потенциала действия, медленный ответ (рис. 6-3 Б), встречается в синоатриальном (SA) узле, области естественного водителя ритма сердца, и в атриовентрикулярном (AV) узле, специализированной ткани, проводящей импульсы сердца из предсердий в желудочки. Сравним эти два принципиальных типа потенциалов действия (рис. 6-3). Потенциал покоя (фаза 4) у быстрого ответа значительно более отрицателен, чем потенциал покоя у медленного ответа. Кроме этого, крутизна нарастания (фаза 0), амплитуда потенциала действия и величина овершута быстрого ответа больше, чем медленного ответа. Амплитуда потенциала действия и крутизна нарастания являются важными факторами, определяющими то, как быстро будет распространяться потенциал действия. В ткани сердца с медленным ответом потенциал действия распространяется более медленно, чем в ткани сердца с быстрым ответом. К тому же проведение возбуждения, скорее всего, будет заблокировано в сердечной ткани с медленным ответом, а не в ткани с быстрым ответом. Медленное проведение и склонность к блоку проведения увеличивает вероятность развития некоторых нарушений ритма. Далее на панелях представлены типичные потенциалы действия, зарегистрированные от клеток SA-узла (B), предсердия (Г) и желудочка (Д). Заметьте, что нумерация фаз потенциала действия дана в соответствии с международными принципами. Рис. 6-3. Изменения трансмембранного потенциала, отводимого от сердечных волокон с быстрым и медленным ответом в изолированной сердечной ткани, помещенной в раствор электролита. А - во время a микроэлектрод был в растворе, который окружает сердечное волокно. Во время b микроэлектрод вошел в клетку. Во время c возник потенциал действия в волокне, в которое введен микроэлектрод. Время от c до d представляет собой фазу абсолютной рефрактерности или эффективный рефрактерный период (ЭРП), а время от d до e представляет относительный рефрактерный период (ОРП). Б - потенциал действия, отводимый от сердечного волокна с медленным ответом. Обратите внимание на то, что по сравнению с волокном, в котором был зарегистрирован быстрый ответ, потенциал покоя медленного волокна менее негативен, фаза 0 (нарастание) потенциала действия менее крутая, амплитуда потенциала действия меньше, фаза 1 отсутствует и ОРП продолжается в значительной мере и в течение фазы 4, после того как волокно полностью реполяризовалось. На панелях В, Г, Д продемонстрированы типичные потенциалы действия, зарегистрированные от клеток СA-узла (B),предсердия (Г) и желудочка (Д). Развертка в B составляет половину от таковой в Г или Д
Типы ионных токов, формирующие фазы ПД рабочего кардиомиоцита Потенциал покоя рабочего кардиомиоцита (Vm) составляет примерно -90 мВ. Когда потенциал покоя рабочего кардиомиоцита внезапно изменяется до порогового уровня, равного примерно -65 мВ, свойства клеточной мембраны резко изменяются. Ионы Na+ входят в кардиомиоциты через существующие в мембране селективные быстрые Na+-каналы. Быстрая деполяризация - фаза 0 - связана с входом ионов Na+ в кардиомиоцит за счет резкого увеличения GNa. По мере того как Na+ стремительно входит в клетку сердца в течение фазы 0, отрицательные заряды внутри клетки нейтрализуются, и Vm становится менее отрицательным. Когда Vmпадает до нуля, электростатическая сила, необходимая для перемещения Na+ в клетку, перестает существовать. Тем не менее пока быстрые Na+-каналы открыты, Na+ продолжает поступать в клетку из-за большого концентрационного градиента. Это пролонгирование входящего Na+ тока приводит к тому, что внутренняя часть клетки становится заряженной положительно. Эта реверсия полярности мембраны и есть так называемый овершут потенциала действия сердца. Такая реверсия электростатического градиента будет, конечно, способствовать ограничению входа дополнительного Na+. Тем не менее на протяжении всего времени, когда направленные внутрь химические силы превосходят направленные вовне электростатические силы, результирующий поток Na+ направлен внутрь, хотя скорость, с которой Na+ поступает в клетку, уменьшается. Поскольку быстрые Na+-каналы являются потенциал-управляемыми, они инактивируются при определенном потенциале и фаза 0 завершится (рис. 6-4). Далее наступает фаза 1 - ранняя реполяризация. Реполяризация происходит быстро вследствие активации транзиторного выходящего тока (Ito) создаваемого главным образом K+. Активация К+-каналов во время фазы 1 вызывает непродолжительный выброс K+ из клетки, потому что внутренняя часть клетки позитивно заряжена и потому, что внутренняя концентрация K+ значительно превосходит внешнюю концентрацию K+. В результате такого транзиторного выхода позитивно заряженных ионов клетка на короткое время частично реполяризуется. Во время фазы 2 - плато потенциала действия - Ca2+ входит в кардиомиоциты через кальциевые каналы, которые активируются и инактивируются гораздо медленнее, чем это делают быстрые Na+-каналы. Во время горизонтального участка фазы 2 такой вход положительного заряда, переносимого Ca2+, сбалансирован выходом положительного заряда, переносимого K+. K+ выходит через каналы, которые проводят в основном Ito, IK и IK1 токи. Как было сказано ранее, ток Ito ответственен за фазу 1, но он не инактивируется полностью до тех пор, пока не закончится фаза 2. Процесс окончательной реполяризации - фаза 3 - начинается в конце фазы 2, когда выход K+ из клетки сердца начинает превышать вход Ca2+. Как мы уже отмечали, по крайней мере, три К+ тока направленных наружу (Ito IK и IK1) вносят вклад в окончательную реполяризацию (фаза 3) клеток сердца (рис. 6-4). В завершение избыток Na+, который входит в клетку в течение фазы 0, активно удаляется Na+/К+-АТФазой. Этот фермент переносит 3 Na+ в обмен на 2 K+, который вышел из клетки главным образом в течение фаз 2 и 3. Точно так же большая часть излишка Ca2+, который вошел в клетку главным образом в течение фазы 2, удаляется преимущественно Na+-Ca2+-обменником, который обменивает 3 Na+ на 1 Ca2+. Восстановление концентраций знаменует фазу 4 - потенциал покоя. Напомним, что IK1 - это inwardly rectifying K+-current, который проводится через К+-кана- лы аномального выпрямления с током входящего направления. Это, inward rectifiers K+-chan- neks - каналы, названные сокращенно Kir (для детализации см. Атлас по физиологии - том 1, рис. 1-16, с. 42 и рис. 1-77, с. 160). Напомним также, что Ito - это fast transient K+-current или transient outward current, который проводится через быстро инактивирующиеся (и поэтому транзиторные) K+-каналы выходящего тока. Это группа потенциал-управляемых K+-каналов - KV. Каналы этого типа обычно образованы членами подсемейств KV1.4, KV3.3, KV3.4, KV4.1 - KV4.3 и вспомогательной β-субъединицы, часто необходимой для феномена быстрой инактивации (для детализации см. Атлас по физиологии - том 1, рис. 1-14, с. 38 и рис. 1-15, с. 40, а также рис. 1-76, с. 158). Напомним, что delayed rectifier K +-channels - K+-каналы задержанного выпрямления, через которые идет ток IK. Это также КV, -каналы, например KV1.1 - KV1.3, KV1.5 - KV1.8, KV2.2, KV3.1, KV3.2 (для детализации см. Атлас по физиологии - том 1, рис. 1-15, с. 40, а также рис. 1-76, с. 158). Рис. 6-4. Ионные токи, текущие через основные ионные каналы, активирующиеся в различные фазы потенциала действия рабочего кардиомиоцита
Ионные токи, формирующие потенциал действия рабочего кардиомиоцита Любой стимул, который делает Vm менее негативным, стремится открыть m-ворота и, таким образом, стремится активировать быстрые Na+-каналы. Величина отдельно взятого потенциала, необходимого, чтобы открыть m-ворота и таким образом активировать Na+-каналы, слегка варьирует от одного канала клеточной мембраны к другому. Когда Vm достигает значений порядка -65 мВ, m-ворота быстрых Na+-каналов стремительно открываются, пока фактически все m-ворота не станут открытыми (фаза 0). Стремительное открытие m-ворот быстрых Na+-каналов ответственно за большое и резкое увеличение Na+ проводимости (GNa), которое происходит в фазу 0. Входящий Na+ ток окончательно останавливается, когда h- (инактивационные) ворота закрываются. Активность h-ворот, как и активность m-ворот, управляется значением Vm. Однако m-ворота открываются очень быстро (примерно за 0,1 м/с), в то время как закрытие h-ворот требует нескольких миллисунд. Фаза 0 заканчивается, когда все h-ворота инактивируются, таким образом, быстрые Na+-каналы закрываются. Такое быстрое закрытие h-ворот вскоре после открытия m-ворот объясняет быстрое возвращение GNa от ее максимума до значения покоя. Затем h-ворота остаются закрытыми до того момента, пока клетка частично реполяризуется во время фазы 3 (рис. 6-5 А1, А2). Фаза 1 - короткий период ограниченной реполяризации происходит быстро вследствие активации транзиторного выходящего тока (ito) создаваемого главным образом K+. Непродолжительный выброс K+ из клетки уменьшает амплитуду потенциала действия. Во время плато - (фаза 2) Ca2+ входит в кардиомиоциты через кальциевые каналы, которые активируются и инактивируются гораздо медленнее, чем это делают быстрые Na+-каналы. Во время фазы 2 такой вход положительного заряда, переносимого Ca2+, сбалансирован выходом положительного заряда, переносимого K+. K+ выходит через каналы, которые проводят в основном Ito, IK и IK1токи. Различные типы Ca2+-каналов были идентифицированы в тканях сердца, но наиболее распространены так называемые, Ca2+-каналы L-типа (long lasting). Открытие Ca2+-каналов приводит к увеличению Ca2+ проводимости (GCa) сразу же после нарастания потенциала действия (рис. 6-5, А2). Во время плато потенциала действия трансмембранный концентрационный градиент K+ фактически такой же, как и во время фазы 4, но Vm положителен, поэтому обе силы, химическая и электростатическая, способствуют выходу K+ из клетки. Если бы GK была во время плато такая же, как во время фазы 4, то выход K+ во время фазы 2 значительно превосходил бы вход Ca2+, и устойчивое плато не возникало бы. Однако по мере того как Vm приближается, а затем достигает положительных значений около пика нарастания потенциала действия, GK внезапно уменьшается. Уменьшенный K+ ток, связанный с понижением GK, предотвращает избыточную потерю K+ из клетки во время плато. Снижение GK как при положительных, так и при небольших отрицательных значениях Vm называется аномальным выпрямлением с током входящего направления (inward rectification). Аномальное выпрямление с током входящего направления присуще нескольким типам K+ токов, включая IK1 ток. Другие K+-каналы - К+-каналы задержанного выпрямления (delayed rectifier channels). Протекающие через них токи обозначаются как IK. Эти каналы закрыты в течение фазы 4, но они активируются потенциалами, которые преобладают к концу фазы 0. Однако активация развивается медленно в течение плато. Следовательно, активация этих каналов ведет к постепенному увеличению GK в течение фазы 2. Таким образом, указанные каналы играют в течение фазы 2 небольшую роль, но вносят определенный вклад в процесс окончательной реполяризации (фаза 3), как это описано ниже. Процесс окончательной реполяризации (фаза 3) начинается в конце фазы 2, когда выход K+ из клетки сердца начинает превышать вход Ca2+. Как мы уже отмечали, по крайней мере, три K+ тока, направленных наружу (Ito IK и IK1), вносят вклад в окончательную реполяризацию (фаза 3) клеток сердца. Необходимо отметить, что определенный вклад вносит ток через катион-неселективные ионные каналы - INS (рис. 6-5 Б). Рис. 6-5. Изменение проводимости мембраны рабочих кардиомиоцитов предсердий и желудочков для ионов (А) и основные ионные токи, принимающие участие в формировании потенциалов действия этих клеток рабочего миокарда (Б). На панели Б в центре - потенциал действия миокардильной клетки желудочка, вверху и внизу - относительная проводимость для входящих и выходящих ионных токов соответственно. Прямые жирные линии для ICl, Ipump и IК(АТФ) указывают лишь на присутствие тока, обусловленного этими каналами или насосами, величины этих токов изменяются при различных физиологических состояниях
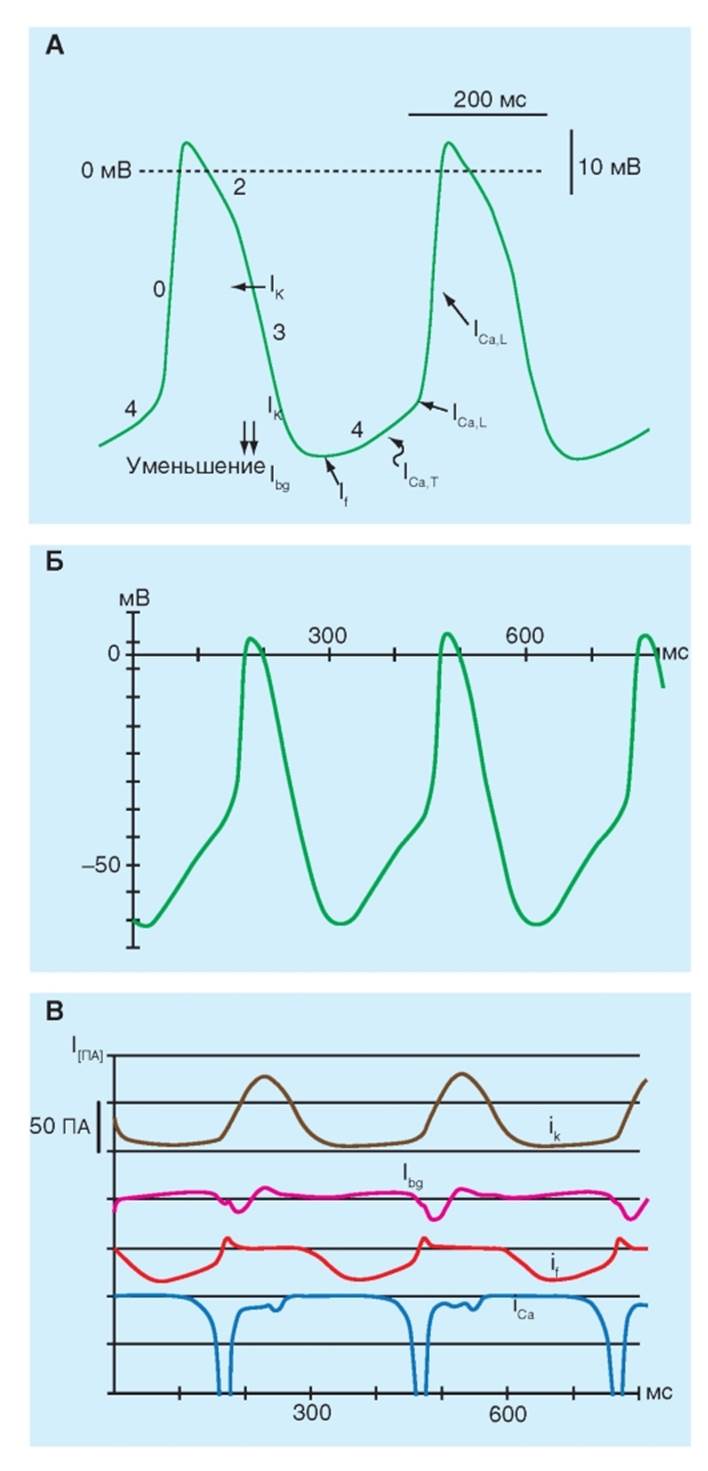
Ионные токи, формирующие потенциал действия пейсмейкерной клетки У медленного же ответа (рис. 6-6 А) нарастание гораздо менее крутое, чем у быстрого. Ранняя реполяризация (фаза 1) отсутствует, плато либо отсутствует, либо значительно меньшей длительности и не такое горизонтальное, а переход от плато к окончательной реполяризации менее выражен. Прежде всего, к клеткам с медленным ответом относятся клетки в синусном (SA) и атриовентрикулярном (AV) узлах. Типичный трансмембранный потенциал действия, зарегистрированный от клетки в SA-узле, изображен на рис. 6-6 А, а реконструированный с помощью компьютерной модели DiFrancescoNoble на рис. 6-6 Б. По сравнению с трансмембранным потенциалом, записанным от кардиомиоцита желудочка, потенциал покоя клетки SA-узла обычно менее негативен, нарастание потенциала действия (фаза 0) менее крутое, плато непродолжительно и реполяризация (фаза 3) более постепенна. Все эти черты характерны для медленного ответа. Нарастание потенциала действия не создается током, направленным внутрь через быстрые Na+-каналы. В таких клетках деполяризация достигается главным образом за счет входа Ca2+ через Ca2+-каналы. Реполяризация в этих волокнах сопровождается инактивацией Ca2+-каналов и увеличенной K+ проводимостью через IK1 и IK каналы (рис. 6-6 В). Трансмембранный потенциал в течение фазы 4 намного менее негативен у клеток, обладающих автоматией в SA- (и AV) узлах, чем у предсердных или желудочковых кардиомиоцитов, потому что IKl (К+-ток аномального выпрямления входящего направления) через К+-каналы аномального выпрямления редко встречается у клеток узлов. Ввиду этого отношение GK к GNa в течение фазы 4 намного меньше у клеток узлов, чем у кардиомиоцитов. Следовательно, во время фазы 4, Vm отклоняется намного больше от равновесного K+потенциала (EK) у клеток узлов, чем у кардиомиоцитов. Однако принципиальное свойство пейсмейкерного волокна, которое отличает его от других рассмотренных нами волокон, заключается в фазе 4. У клеток, не обладающих автоматией, мембранный потенциал остается постоянным в течение этой фазы, в то время как пейсмейкерное волокно характеризуется медленной диастолической деполяризацией в течение фазы 4. Деполяризация развивается с постоянной скоростью до достижения порога, запуская потенциал действия. Рис. 6-6. Основные ионные токи, активирующиеся в различные фазы потенциала действия пейсмейкерной клетки в синоатриальном узле. А - потенциалы действия и ионные токи, участвующие в формировании потенциалов действия пейсмейкерной клетки в синоатриальном узле. IK - калиевый ток задержанного выпрямления, Ibg - фоновый ток утечки, If - пейсмейкерный ток, активируемый гиперполяризацией, ICa,T - T-тип (от transient - преходящий) Са2+-токов активируется слабой деполяризацией и эти токи мимолетные (преходящие), ICa,L (от long-lasting - долгодлящийся) Са2+-токов, требующий сильной деполяризации для активации; он долго-длящийся и блокируется органическими антагонистами L-типа Са2+-каналов, включая дигидропиридины, фенилалкиламины и бензотиазепины. Б - реконструкция спонтанной активности клетки синоатриального узла с помощью компьютерной модели DiFrancesco-Noble. Показаны потенциалы действия. В - реконструкция ионных токов клетки синоатриального узла с помощью компьютерной модели DiFrancesco-Noble. Показаны амплитуды ионных токов IK, Ibg, If и ICa(ICa,L + ICa,T)
Ионные механизмы автоматии Несколько ионных токов вносят вклад в медленную диастолическую деполяризацию, которая характерна для клеток сердца, обладающих автоматией. В пейсмейкерных клетках SA-узла, по крайней мере, три ионных тока опосредуют медленную диастолическую деполяризацию: (1) входящий ток, If, вызванный гиперполяризацией; (2) входящий Ca2+ ток, ICa и (3) выходящий K+ ток, IK (рис. 6-7 А и Б). Входящий ток If (funny) активируется ближе к концу реполяризации. Этот «странный» ток переносится главным образом Na+ через специфические каналы, которые отличаются от быстрых №+-каналов. Току был пожалован титул «странного», потому что его первооткрыватели не ожидали обнаружить направленный внутрь Na+ ток в пейсмейкерных клетках после завершения реполяризации. Этот ток активируется по мере того, как мембранный потенциал становится более негативным, чем приблизительно -50 мВ. Чем более негативен мембранный потенциал в это время, тем больше активация If тока. Второй ток, ответственный за диастолическую деполяризацию, Ca 2+ ток, ICa. Этот ток активируется к концу фазы 4 по мере того, как трансмембранный потенциал достигает величины около - 55 мВ. Как только каналы активированы, вход Ca2+ в клетку увеличивается. Этот приток ускоря- ет диастолическую деполяризацию, которая затем приводит к фазе нарастания потенциала действия. Уменьшение концентрации внешнего Ca2+ или добавление антагонистов кальциевого канала уменьшает амплитуду потенциала действия и крутизну медленной диастолической деполяризации в клетках SA-узла. Прогрессивной диастолической деполяризации, опосредованной двумя входящими токами, If и ICa, противодействует выходящий ток K+- ток задержанного аномального выпрямления IK. Эта утечка K+ стремится реполяризовать клетку после нарастания потенциала действия. K+ продолжает выходить наружу в течение значительного времени после максимальной реполяризации, но этот выход уменьшается на всем протяжении фазы 4. Как только ток уменьшается, его противодействие деполяризующим влияниям двух входящих токов (ICa и If) также постепенно уменьшается. Ионные основы автоматии в пейсмейкерных клетках AV-узла похожи на таковые в клетках SA-узла. Подобные механизмы также отвечают за автоматию в волокнах Пуркинье желудочков, за исключением того, что в них не участвует Ca2+ ток. Другими словами, медленная диастолическая деполяризация опосредована преимущественно неустойчивостью между влиянием вызванного гиперполяризацией входящего тока If и постепенно уменьшающимся выходящим K+ током IK. Рис. 6-7. Изменение проводимости мембраны пейсмейкерных клеток синоатриального узла для ионов (А) и основные ионные токи, принимающие участие в формировании потенциалов действия этих клеток (Б). На панели Б в центре - потенциал действия клетки синоатриального узла, вверху и внизу - входящие и выходящие ионные токи соответственно. Прямые жирные линии для Ipump и Ibg указывают лишь на присутствие тока, обусловленного этими каналами или насосами, величины этих токов изменяются при различных физиологических состояниях. Знак вопроса указывает на то, что роль этих токов в формировании потенциала действия синоатриальных клеток до конца не выяснена, тогда как знак «0» говорит о том, что этот ток отсутствует
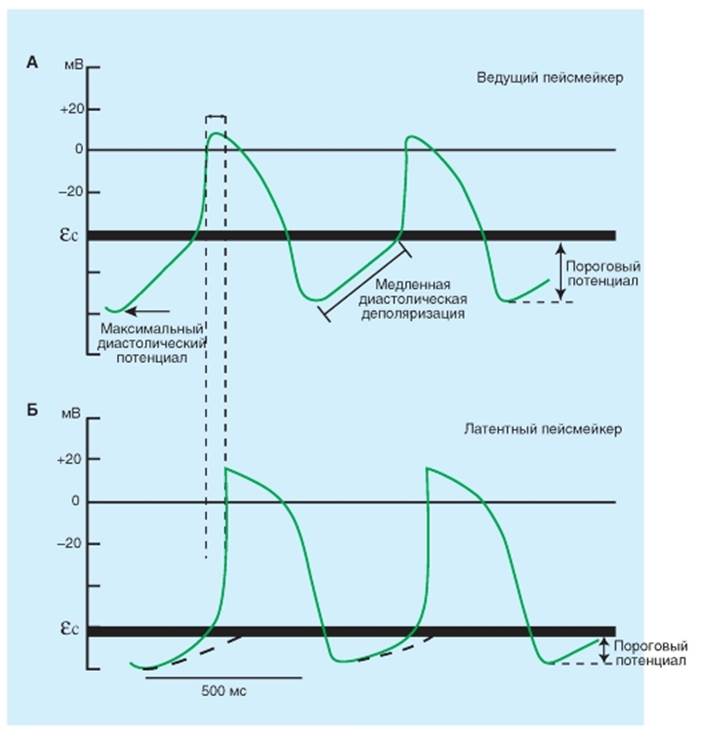
Истинные и латентные пейсмейкерные комплексы Свойство автоматии - ведущей пейсмейкерной активности - позволяет сердцу сокращаться даже тогда, когда сердце изолировано из организма. Если коронарные сосуды извлеченного сердца искусственно перфузировать раствором электролита, насыщенного кислородом, ритмические сердечные сокращения сохраняются в течение многих часов. По крайней мере, некоторые клетки предсердий и желудочков могут возбуждаться и инициировать сокращения. Такие клетки главным образом находятся в тканях узлов или специализированных проводящих волокнах сердца. Областью сердца млекопитающих, которая обычно генерирует импульсы с самой большой частотой (рис. 6-8 А), является синоатриальный (SA) узел. Это главный водитель ритма сердца. Детализированное картирование электрических потенциалов на поверхности правого предсердия показывает, что существуют две или три области автоматии, находящиеся в 1 или 2 см от SA-узла, и составляют совместно с SA-узлом предсердный пейсмейкерный комплекс. Иногда все эти локусы одновременно инициируют импульсы. Однако принципиальное свойство пейсмейкерного волокна, которое отличает его от других рассмотренных нами волокон, заключается в фазе 4. У клеток, не обладающих автоматией, потенциал остается постоянным в течение этой фазы, в то время как пейсмейкерное волокно характеризуется медленной диастолической деполяризацией в течение фазы 4. Деполяризация развивается с постоянной скоростью до достижения порога, запуская потенциал действия. Однако существуют и латентные пейсмейкеры. Когда SA-узел или другие составляющие предсердного пейсмейкерного комплекса удалены или разрушены, клетки водителя ритма AV-узла обычно берут на себя пейсмейкерную функцию всего сердца. Через некоторое время клетки предсердий, обладающие автоматией, обычно становятся доминирующими. Волокна Пуркинье представляют специализированную проводящую систему желудочков, которая также способна проявлять автоматию. Как правило, эти волокна разряжаются с очень низкой частотой. Когда AV-узел не может проводить импульсы от предсердий к желудочкам (рис. 6-8 Б), собственный желудочковый водитель ритма в сети волокон Пуркинье инициируют сокращения желудочков, но только с частотой от 30 до 40 ударов в минуту. Рис. 6-8. Потенциалы действия ведущего пейсмейкера. А - клетки синусового узла и латентного пейсмейкера. Б - клетки пучка Гиса. Вертикальными пунктирными линиями отмечены моменты начала возбуждения пейсмейкеров, стрелкой отмечено время проведения возбуждения от истинного пейсмейкера к латентному. Пунктирными линиями показано время достижения порогового потенциала латентными пейсмейкерными клетками. В результате они возбуждаются в ритме, заданном истинным пейсмейкером, поскольку в латентных пейсмейкерах медленная диастолическая деполяризация не успевает достигнуть порогового уровня. ɛс - критический уровень деполяризации
Возможные пути изменения частоты разрядов пейсмейкерных клеток Частота разрядов пейсмейкерных клеток, определяющих частоту сердечных сокращений, может варьировать в результате изменения следующих условий: (1) при изменении скорости диастолической деполяризации в фазу 4; (2) при изменении максимально отрицательной величины мембранного потенциала в фазу 4; (3) при изменении порогового потенциала за счет сдвига величины критического уровня деполяризации; (4) при изменении скорости нарастания переднего фронта деполяризации и скорости реполяризации (рис. 6-9). Когда скорость медленной диастолической деполяризации увеличивается (от 2 до 1 на рис. 6-9 А1), потенциал порога достигается раньше и частота сердечных сокращений увеличивается. Повышение порогового потенциала (от Ес-1 до Ес-2 на рис. 6-9 А2) задерживает начало фазы 0 (от момента времени 2 до момента времени 3) и частота сердечных сокращений соответственно уменьшается. Точно так же, когда максимальный, негативный потенциал увеличен (от 1 до 4 на рис. 6-9 А2), то требуется больше времени, чтобы достигнуть порога Ес-2, когда крутизна фазы 4 остается неизменной, а частота сердечных сокращений, следовательно, уменьшается. Рис. 6-9. Основные пути, обусловливающие изменения частоты возбуждения пейсмейкерных клеток. А - причины изменения частоты пейсмейкерной активности. А1 - изменение частоты спонтанной активности за счет изменения скорости медленной диастолической деполяризации (1 и 2). А2 - изменение частоты спонтанной активности за счет изменения порогового уровня деполяризации (Ес-1, Ес-2 - 1 и 4). Изменение частоты спонтанной активности за счет изменения уровня максимального диастолического потенциала (2, 3 и 5). Б - влияние кратковременного раздражения блуждающего нерва (стрелка) на трансмембранный потенциал, регистрируемый от пейсмейкерной клетки SA-узла препарата изолированного предсердия кошки. Длительности сердечного цикла, в миллисундах, обозначены числами наверху рисунка
Влияние величины мембранного потенциала покоя на скорость распространения потенциала действия Уровень потенциала покоя мембраны управляет скоростью проведения потенциалов действия, поскольку влияет на амплитуду потенциала действия и крутизну его нарастания. Скорость распространения потенциала действия по миокарду определяют следующие факторы: (1) величина мембранного потенциала покоя, (2) амплитуда ПД и (3) скорость нарастания его переднего фронта в фазу 0. (1) Чем более деполяризована мембрана, тем меньше скорость распространения независимо от причин возникновения. В основе этого явления лежит инактивация потенциал-управляемых h-ворот. А именно, когда начинается регенеративная деполяризация, то часть быстрых натриевых каналов могут быть уже инактивированы. (2) Амплитуда ПД определяет разность потенциалов между возбужденным и невозбужденным участком, т.е. величину локальных токов, которые деполяризуют мембрану до порогового уровня. (3) Скорость нарастания переднего фронта влияет на величину разности потенциалов между возбужденным и еще не возбужденным участками. Иными словами, если ПД развивается медленно, то это сопровождается снижением возникающей разности потенциалов, поскольку покоящаяся часть деполяризуется постепенно по отношению к активной зоне. Этими механизмами можно объяснить более медленное распространение возбуждения потенциалов с медленным ответом. Следует учитывать, что блокируются в первую очередь потенциалы с медленным ответом. Трансмембранный потенциал может изменяться в сторону деполяризации по следующим причинам: (1) изменилась внеклеточная концентрация K+; (2) у волокон сердца, способных к автоматии, Vm постепенно становится менее негативным во время фазы 4; и (3) при преждевременном возбуждении мембрана клетки полностью не реполяризуется после предыдущего возбуждения. Вообще говоря, чем менее отрицателен уровень Vm, тем меньше скорость распространения импульса, независимо от причины изменения Vm. Уровень Vm влияет на скорость проведения, потому что инактивация h-ворот быстрых Na+-каналов, является потенциал-управляемой. Чем менее отрицателен Vm, тем больше количество h-ворот, которые стремятся закрыться. В течение нормального процесса возбуждения, деполяризация развивается так быстро в течение фазы 0, что сравнительно медленные h-ворота не закрываются до конца этой фазы. Однако если неполная деполяризация произведена постепенно, например, как при повышении уровня внешнего K+, то ворота имеют вполне достаточное время, чтобы закрыться и таким образом инактивировать некоторые из Na+-каналов. Когда клетка не полностью деполяризована, многие из Na+-каналов уже инактивированы; таким образом, только часть Na+-каналов способна проводить направленный внутрь Na+ ток в течение фазы 0. На рис. 6-10 показаны результаты эксперимента, в котором потенциал покоя пучка волокон Пуркинье уменьшали, изменяя величину [K+]o. Ткань стимулируется на некотором удалении от клетки с вживленным электродом, и артефакт от раздражения (St) появляется как двухфазное отклонение перед фазой 0. Интервал от этого артефакта до начала фазы 0 обратно пропорционален скорости проведения. Когда [K+]o равна 3 мМ (A и Е), Vm составляет -82 мВ, и наблюдается крутой наклон в фазе 0. В конце фазы 0, овершут достигает 30 мВ. Следовательно, амплитуда потенциала действия составляет 112 мВ. Когда [K+]o постепенно увеличивается до 16 мМ (от Б к Д), потенциал покоя постепенно становится менее негативным. В то же самое время и амплитуды, и длительности потенциалов действия, и крутизны нарастания уменьшаются. Как следствие, скорость проведения прогрессивно снижается. При уровнях [K+]o в 14 и 16 мМ (Д и E), Vm достигает значений, достаточных, чтобы инактивировать все быстрые Na+-каналы. Потенциалы действия на рис. 6-9 Д и E являются характерными медленными ответами. Рис. 6-10. Зависимость мембранного потенциала и потенциала действия клеток Пуркинье от внеклеточной концентрации калия (St - стимуляция)
Взаимосвязь частоты стимуляции и длительности потенциала действия кардиомицита Изменения частоты стимулирующих импульсов изменяют длительность потенциалов действия в клетках сердца (рис. 6-11) и таким образом изменяют их рефрактерные периоды. Следовательно, изменения частоты стимулирующих импульсов часто являются важными факторами при инициировании или прекращении некоторых аритмий. Изменения длительности потенциала действия, которые происходили при уменьшении длительности межстимуляционного интервала от 2000 до 200 мс в волокне Пуркинье показаны на рис. 6-11. Обратите внимание, что по мере того, как длительность межстимуляционного интервала сокращается, длительность потенциала действия уменьшается. Прямая корреляция между длительностью потенциала действия и длительностью межстимуляционного интервала опосредована изменениями в GK, во что вовлечены, по крайней мере, два типа K+-каналов, а именно, K+-каналы задержанного выпрямления (delayed rectifier channels), формирующие ток IK и быстро инактивирующиеся (и поэтому транзиторные) K+-каналы, которые проводят направленный наружу Ito. Ток IK активируется при значениях Vm близких к нулю, но ток активируется медленно и остается активированным сотни миллисунд. Ток IK также очень медленно инактивируется. Следовательно, поскольку длительность межстимуляционного интервала уменьшается, каждый следующий потенциал действия имеет тенденцию возникать в начале периода инактивации тока IK предшествующего потенциала действия. Таким образом, чем короче длительность межстимуляционного интервала, тем больше направленный наружу K+-ток в течение фазы 2, и, следовательно, короче длительность потенциала действия. Ток Ito влияет на связь между частотой стимуляции и длительностью потенциала действия. Ток Ito также активируется при значениях потенциалов, близких к нулю, и его величина изменяется обратно пропорционально длине сердечного цикла. Следовательно, при уменьшении межстимуляционного интервала, результирующее увеличение направленного наружу K+ тока сокращает плато. Относительные вклады IK и Ito во взаимосвязь между длительностью потенциала действия и длиной сердечного цикла варьируют в сердцах разных видов животных. Рис. 6-11. Влияние длительности межстимуляционного интервала (CL) на длительность потенциала действия (APD) клеток Пуркинье
Влияние стимуляции вегетативных нервов на форму потенциалов клеток синусного, атриовентрикулярного узлов и кардиомиоцитов предсердий Обычно частота разряда водителя ритма регулируется действием обоих отделов вегетативной нервной системы. Усиленная симпатическая нервная активность, посредством высвобождения норадреналина, повышает частоту сердечных сокращений, увеличивая, преимущественно, крутизну медленной диастолической деполяризации рис. 6-12 А1, А2, А3). Этот механизм увеличения частоты сердечных сокращений осуществляется при физической нагрузке, беспокойстве, или при некоторых заболеваниях. Повышенная активность блуждающего нерва, посредством высвобождения ацетилхолина, уменьшает частоту сердечных сокращений, гиперполяри- зуя клеточную мембрану пейсмейкера и уменьшая крутизну медленной диастолической деполяризации (рис. 6-12 Б1, Б2, Б3). Эти механизмы уменьшения частоты сердечных сокращений осуществляются, когда преобладает действие блуждающего нерва. Примером крайнего случая является вазовагальный обморок: короткий период головокружения или потери сознания, вызванного интенсивным всплеском вагусной активности. Этот тип обморока является рефлекторным ответом на боль или некоторые психологические стимулы. Изменения активности внутрисердечной нервной системы обычно не изменяют частоту сердечных сокращений посредством изменения порогового значения Vm, что инициирует разряд пейсмейкерной клетки узла. Однако некоторые антиаритмические препараты, например, хинидин и прокаинамид, поднимают пороговый потенциал клеток, обладающих автоматией до менее негативных величин за счет смещения ɛс. Рис. 6-12. Влияния парасимпатической и симпатической стимуляции на форму потенциалов действия клеток синоатриального узла (А1, Б1), атриовентрикулярного узла (А2, Б2) и рабочего миокарда предсердий (А3, Б3). На «В», также приведены кривые изометрических сокращений предсердий.
|
|||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-20; просмотров: 570; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.117.145.67 (0.017 с.) |